- Преподавателю
- Биология
- СПЕШИТЕ СПАСТИ ПЛАНЕТУ ! Методическое пособие к видеосериалу Часть 2
СПЕШИТЕ СПАСТИ ПЛАНЕТУ ! Методическое пособие к видеосериалу Часть 2
| Раздел | Биология |
| Класс | 10 класс |
| Тип | Другие методич. материалы |
| Автор | Ганюшин Л.А. |
| Дата | 08.05.2015 |
| Формат | doc |
| Изображения | Есть |

Научно-методический центр по эколого-валеологическому образованию
и воспитанию населения
СПЕШИТЕ СПАСТИ ПЛАНЕТУ !
Методическое пособие к видеосериалу
Часть 2
Серия: Экология Э-3
Переработал и дополнил
Ганюшин Л.А.
Новомосковск, 2000 г.
Настоящей работой мы продолжаем серию методических материалов, посвященных экологическим проблемам, истории экологии и охраны природы, толкованию законов экологии и экологического права.
Оглавление
Понимание экосистем
Учебные цели
Рекомендуемая литература
Обзор материала
Дополнительные материалы
Вопросы для обсуждения или письменных заданий
Практические задания
Контрольные вопросы
Библиография
Учебные цели
После изучения рекомендованной литературы каждый учащийся, студент должен уметь:
- различать роли, которые играют продуценты, консументы и редуценты в первичных экосистемах;
- составить диаграмму и описать, по крайней мере, два биогеохимических цикла и обсудить основные аспекты воздействия человека на каждый из них;
- определить и обсудить важность фотосинтеза как в наземных, так и в водных экосистемах;
- объяснить, как трофическая структура природных экосистем иллюстрирует второй закон термодинамики - закон энтропии;
- перечислить пять факторов, влияющих на климат Земли, определить и обсудить факторы, на которые оказывает влияние деятельность человека;
- описать три основных типа экосистем суши;
- объяснить, как температура и количество осадков определяют тип наземных экосистем;
- описать два основных типа водных экосистем;
- объяснить роль солености в определении типа водных экосистем;
- перечислить главные различия между "умеренным" и "тропическим" вариантами трех типов наземных экосистем.
Рекомендуемая литература
- Миллер Тайлер Жизнь в окружающей среде: Программа всеобщего экологического образования, В 3-х кн., Изд. группа Прогресс, Пангея, М., 1993
Глава 4. Экосистемы: что они представляют и как работают?
4.1. Системы поддержания жизни на Земле: общий обзор
4.2. Экосистемы: типы и составляющие
4.3. Потоки энергии в экосистемах
4.4. Круговорот вещества в экосистемах
Глава 5. Основные типы экосистем
5.1. Климат: краткая характеристика
5.2. Основные типы экосистем суши: пустынные, травянистые и лесные
5.3. Водные экосистемы
Глава 6.
6.1. Реакция живых систем на влияние окружающей среды
6.2. Реакция популяций на резкие изменения окружающей среды
6.3. Реакция сообществ и экосистем на стресс
6.4. Воздействие человека на экосистемы
- Небел Б. Наука об окружающей среде: Как устроен мир, В 2-х томах, М., Мир, 1993
Часть I
Что такое экосистемы и как они функционируют
Глава 1. Экосистемы - что это такое?
Определение и примеры экосистем
Структура экосистем
Почему в разных регионах сформировались разные экосистемы?
Экология и человек
Леса умеренного пояса
Степи
Пустыни
Хвойные леса (тайга)
Тундра
Саванны
Дождевые тропические леса
Глава 2. Экосистемы: как они функционируют?
Элементы, жизнь, организация и энергия
Принципы функционирования экосистем
Связь с жизнью человека
Глава 3.
Экосистемы: почему они устойчивы? Почему изменяются?
Главное - равновесие
Связь с жизнью человека
Глава 4. Экосистемы: адаптация и изменения или вымирание
Как виды изменяются и адаптируются и что им мешает это делать
Связь с жизнью человека
- Мамедов Н.М., Суравегина И.Т. Экология: Учебное пособие, 9-11 классы, М., Школа-Пресс, 1996
Глава 2. Экосистема - основной элемент биосферы
2.1. Особенности экосистемы
2.2. Трофическая структура сообщества
2.6. Самоочищение экосистемы
2.7. Развитие экосистемы
2.8. Реакции экосистем на загрязнение и разрушение
- Миркин Б.М., Наумова Л.Г. Экология России, 9-11 классы, М., АО МДС, 1996
Глава 2. Экосистема
Глава 3. Экологическое равновесие и сукцессии естественных экосистем
Глава 4. Естественные экосистемы России и их использование
Глава 5. Структура и устойчивость социоприродных экосистем
- Криксунов Е.А., Пасечник В.В., Сидорин А.П. Экология, 9 класс, Изд. дом Дрофа, 1995
4.1. Сообщество, экосистема, биоценоз
4.6. Живые организмы и круговорот веществ в экосистеме
- Алексеев С.В. Экология, Учебник для 9 класса, СПб., СМИО ПРЕСС, 1997
Глава 3. Экология сообществ (синэкология)
- Чернова Н.М., Галушин В.М., Константинов В.М. Основы экологии, Учебник для 9 класса, М., Просвещение, 1997
Глава 3. Экосистемы
- Демина Т.А. Экология, природопользование, охрана окружающей среды, М., Аспект Пресс, 1996
Глава 2. Закономерности развития биосферы
Обзор материала
Эта часть работы является подготовительной к видеосериалу и в ней представлены основные положения экологии, учения о том, как виды растений, животных и микроорганизмов взаимодействуют между собой, а также как влияют на них химические и физические факторы, действующие в среде их обитания.
Слово "экология" происходит от греческих слов oikos (жилище) и logos (учение). Таким образом, экология является наукой о нашем доме, нашей планете, которая обеспечивает средствами существования как Homo Sapiens, так и другие населяющие ее виды.
В рекомендуемой литературе в соответствующих главах дается определение экосистем и рассматриваются механизмы их действия. Представлены два фундаментальных процесса, общие для всех экосистем: поглощение солнечной энергии и ее прохождение через экосистему (в соответствии со вторым законом энергии), а также накопление и круговорот питательных веществ в биогеохимических циклах.
В пособии рассматриваются ограничивающие факторы и пределы толерантности, которые определяют численность и распределение растительных и животных видов, а также различные варианты сосуществования видов в экосистемах; описываются основные типы экосистем суши и водных экосистем, а также рассматриваются влияние климата (средней температуры и осадков) на наземные экосистемы и влияние концентрации растворенных в воде солей (солености) на водные экосистемы.
Каждая точка на Земле является частью экосистемы. Каждый наш вдох, любая съеденная пища и каждый глоток воды напрямую связывают нас с биогеохимическими циклами. Как будет ясно из десяти телепрограмм курса "Спешите спасти планету", коллективное воздействие человечества на природу достигло таких масштабов, когда деятельность людей влияет и на поступление солнечной энергии и круговорот энергии в экосистемах, на многообразие видов растений и животных, составляющих эти экосистемы, и на общий круговорот газов, минеральных элементов и воды.
Понимание экосистем и того, как они функционируют, является первым шагом к пониманию влияния, которое человечество оказывает на окружающую среду, и далее, к пониманию того, как мы можем привести желания и потребности людей в соответствие с границами, установленными продуктивностью и способностью биосферы к восстановлению.
Дополнительные материалы
Авторские лекции
Экосистема - ключевое понятие экологии
Из определения "Экология" следует, что наука рассматривает взаимодействие живых организмов и костного вещества в их единстве и развитии.
Такое взаимодействие определенным образом организовано и характерно для любой экологической системы.
Экосистему обычно определяют как совокупность различных видов растений, животных и микроорганизмов, взаимодействующих друг с другом и с окружающей их средой посредством обмена веществом, энергией и информацией таким образом, что вся эта система сохраняется и остается устойчивой в течение продолжительного времени (от нескольких лет - агроэкосистемы, до миллиардов лет - биосфера). Все экосистемы взаимосвязаны и образуют в своей совокупности единое целое, глобальную экосистему - биосферу.
Из этого определения можно сделать два основных вывода:
-
любая экосистема обязательно представляет собой совокупность живых (биотических) и неживых (абиотических) компонентов;
-
устойчивость экосистемы обусловлена наличием определенных обменных процессов, реализация которых возможна лишь при определенной структуре биотических и абиотических компонентов и их взаимосвязи.
Необходимо подчеркнуть, что любая совокупность (сообщество) живых организмов характерна для экосистемы, а лишь определенным образом структурированная.
Структура и иерархия экосистем
Мир основан на единстве живого и неживого. Суть этого единства состоит в следующем. Из элементов неживой природы, главным образом молекул СО2 и Н2О, под воздействием энергии солнца синтезируются органические вещества, составляющие все живое на планете.
Процесс создания органического вещества в природе протекает одновременно с противоположными процессами - потребление и разложение этого вещества вновь на исходные неорганические соединения. Все эти процессы протекают в рамках экосистем различных уровней иерархии. Замкнутость обменных процессов вещества и обуславливает определенную биотическую структуру экосистем.
Движущей силой в любой материальной системе служит энергия. В экосистемы она поступает главным образом от Солнца. Это энергия в виде излучений. Эта энергия воспринимается растениями. За счет содержащегося в них пигмента хлорофилла растения улавливают энергию излучения Солнца и используют ее для синтеза основы основ органического вещества - глюкозы С6Н12О6. Для этого они извлекают атомы углерода и кислорода из молекул СО2, поглощаемого ими из воздуха, и атомы водорода и кислорода - из содержащихся в нем молекул воды. Это и есть процесс фотосинтеза:
6СО2 + 6Н2О солнечный свет = глюкоза С6Н12О6 +6О2
Излишек атомов О2 выделяется в атмосферу в виде газообразного кислорода.
Кинетическая энергия солнечного излучения преобразуется, таким образом, в потенциальную энергию, запасенную глюкозой. Из глюкозы вместе с получаемыми из почвы минеральными элементами питания (биогенами) образуются все ткани растительного мира - белки, углеводы, жиры, липиды, ДНК, РНК, т.е. органическое вещество планеты.
Кроме растений продуцировать органическое вещество могут некоторые бактерии. Они создают свои ткани, запасая в них, как и растения, потенциальную энергию, из углекислого газа без участия солнечной энергии. Вместо нее они используют энергию, которая образуется при окислении неорганических соединений, например, аммиака, железа и особенно серы (в глубоких океанических впадинах, куда не проникает свет, но где в изобилии есть Н2S, обнаружены уникальные экосистемы). Это так называемая энергия химического синтеза, поэтому такие организмы называют хемосинтетиками.
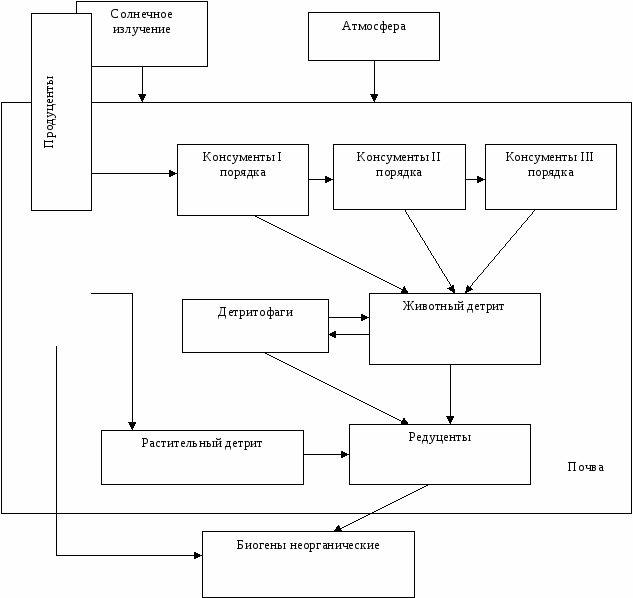
Таким образом, растения и хемосинтетики создают органическое вещество из неорганических составляющих с помощью энергии окружающей среды. Их называют продуцентами или автотрофами. Высвобождение запасенной продуцентами потенциальной энергии обеспечивает возможность существования всех остальных видов живого на планете. Виды, потребляющие созданную продуцентами органику как источник вещества энергии для своей жизнедеятельности, называют консументами или гетеротрофами.
Консументы - это самые разнообразные организмы от микроскопических бактерий до синих китов: это простейшие, насекомые, пресмыкающиеся, рыбы, птицы и, наконец, млекопитающие, включая человека.
Консументы, в свою очередь, подразделяются на ряд подгрупп в соответствии с различиями в источниках их питания.
Животные, питающиеся непосредственно продуцентами, называются первичными консументами (или консументами I порядка). Их самих употребляют в пищу вторичные консументы. Например, кролик, питающийся морковкой, - это консумент первого порядка, а лиса, охотящаяся за кроликом - консумент второго порядка. Бывают консументы третьего и крайне редко четвертого порядков. Причем некоторые виды живых организмов соответствуют нескольким таким уровням. Например, когда человек ест овощи - он консумент первого порядка, питаясь говядиной - вторичный консумент, а если он ест хищную рыбу, то выступает в роли консумента третьего порядка.
Первичные консументы, питающиеся только растениями, называются растительноядными или фитофагами. Консументы второго и более высоких порядков - плотоядные. Виды, с равным успехом употребляющие в пищу как растения, так и животных, относятся к всеядным (медведь, человек).
Мертвые растительные животные остатки, например, опавшие листья, трупы животных, продукты систем выделения и т.д. - называются детритом. Это органика! Существует множество специализированных на питании детритом организмов. Они называются детритофагами. Примером могут служить грифы, шакалы, черви, раки, термиты, муравьи и т.п. Как и в случае обычных консументов, различают первичных детритофагов, питающихся непосредственно детритом, вторичных и т.д.
Наконец, значительная часть детрита в экосистеме, в частности, опавшие листья, валежная древесина в своем исходном виде не поедается животными, а гниет и разлагается в процессе питания им грибов и бактерий.
Поскольку грибы и бактерии столь специфичны, их обычно выделяют в особую группу детритофагов и называют редуцентами. Редуценты служат на Земле санитарами и замыкают биогеохимический круговорот веществ, разлагая органику на исходные неорганические составляющие (СО2, воду и т.п.).
Таким образом, несмотря на многообразие экосистем, все они обладают структурным сходством. В каждой из них можно выделить фотосинтезирующие растения - продуценты, различные уровни консументов, детритофагов и редуцентов. Они и составляют биотическую структуру экосистем.
Получив представление о структуре экоситем уточним следующие понятия.
Вид - множество организмов, сходных по строению и (или) способных скрещиваться друг с другом, давая плодовитое потомство.
Популяция - группа в пределах вида, особи которой скрещиваются друг с другом и размещаются на отдельной территории.
Сообщество - это совокупность совместно проживающих популяций разных видов в пределах какого-либо естественного пространства.
Могут рассматриваться отдельно сообщество растений - фитоценоз ("ценоз" - сообщество), сообщество животных - зооценоз, сообщество микроорганизмов - микробоценоз, а могут - в различных комбинациях (для понятия "сообщество" неважно ни местонахождение, ни внешние условия, ни выполняемые функции, это обобщенное название различных сочетаний популяций живых организмов).
Совокупность живых организмов, включающая все биотические компоненты (продуценты, консументы, детритофаги, редуценты) и населяющая определенный ограниченный в пространстве участок с однородными абиотическими условиями (биотоп), называется биоценозом. Экосистемы различного уровня организации функционально соподчинены, т.е. имеют определенную иерархическую структуру. Смысл иерархии состоит в том, что более простые системы объединяясь создают более сложные с новыми функциями. Более высокий уровень организации - это не просто количественное сложение более простых структур. Главное, что каждый последующий иерархический уровень сохраняет свойства и законы, действующие на уровнях с более низкой организацией и плюс приобретает новые. Новые (эмерджентные) свойства в более сложно организованных системах появляются из-за возникновения в них новых функциональных связей. Экосистема каждого уровня организации имеет свой биотический круговорот. Высший уровень иерархии занимает глобальная экосистема - биосфера.
Биотическая структура экосистем
Поток энергии в экосистемах
Жизнь как термодинамический процесс
Чтобы понять благодаря каким механизмам экосистемы поддерживают свою структуру и функционируют необходимо разобраться с процессами энергомассообмена, происходящими в них.
Закон сохранения энергии и закон энтропии - это фундаментальные законы природы. Они справедливы и для экосистем. Все разнообразие проявления жизни сопровождается превращениями энергии, хотя при этом энергия не создается и не уничтожается (I закон термодинамики). Энергия, получаемая в виде солнечной излучения биосферой Земли, уравновешивается энергией, излучаемой с поверхности Земли в форме невидимого теплового излучения. Сущность жизни состоит в непрерывной последовательности таких изменений как рост самовоспроизведения и синтез сложных химических соединений. Без переноса энергии, сопровождающего все эти изменения, не было бы ни жизни, ни экосистем, в том числе биосферы. Наша планета и цивилизация - лишь одно из замечательных явлений природы, зависящих от постоянного притока концентрированной энергии солнечного излучения.
Земля получает ~ 510-10 доли общей энергии, излучаемой Солнцем, это ~ 1017 Вт или 51024 Дж/год. Из этого количества энергии 33 % отражается облаками и поверхностью суши, а также тонкодисперсной пылью в верхних слоях атмосферы. Эта часть составляет альбедо Земли. 67 % энергии поглощается атмосферой и земной поверхностью и после ряда превращений уходит в космическое пространство. По оценкам ученых всего лишь 1 % солнечной энергии, падающей на Землю, принимает участие в фотосинтезе. Такой низкий КПД фотосинтеза некоторые объясняют малой концентрацией СО2 в атмосфере и гидросфере. Из этого количества энергии, поглощаемой земными растениями, лишь очень небольшая часть энергии превращается в химическую энергию пищи. Весь остальной живой мир получает необходимую энергию из органических веществ, созданных фотосинтезирующими растениями или хемосинтезирующими микроорганизмами.
Перенос энергии пищи от ее источника - автотрофа (растений) через ряд организмов, называется пищевой (трофической) цепью. Основная часть потребляемой в ней энергии идет на поддержание жизнедеятельности, т.е. на процесс расщепления органических молекул с выделением энергии, называемый клеточным дыханием. В целом он противоположен фотосинтезу: в присутствии кислорода происходит разрушение глюкозы по уравнению
С6Н12О6 + 6О2 = 6СО2 + 6Н2О + % энергия
И лишь сравнительно небольшая часть потребляемой с пищей энергии идет на построение тела, рост и размножение.
Итак, на каждом этапе передачи энергии от одного организма к другому значительная часть ее превращается в тепло и рассеивается в соответствии со II законом термодинамики (законом энтропии). Это самопроизвольный процесс.
В неживой природе самопроизвольные процессы ведут систему к состоянию равновесия. Если живую неуравновешенную с окружающей средой систему изолировать, то всякое движение в ней скоро прекратится. В результате трения, теплопроводности, химических реакций и других самопроизвольных процессов, потенциалы (температура, концентрация и т.п.) выравниваются, система в целом угаснет и превратится в инертную массу, находящуюся в состоянии термодинамического равновесия с окружающей средой, т.е. с максимальной энтропией. Это наиболее вероятное для такой системы состояние и самопроизвольно без внешних воздействий она уже выйти из него не сможет.
У живых систем есть одно принципиальное отличие от неживых - они совершают постоянную работу против уравновешивания с окружающей средой. Советский ученый Бауэр назвал это "всеобщим законом биологии", который имеет ясный термодинамический смысл: как в неживых системах устойчиво их равновесное состояние, так в живых системах устойчиво неравновесное состояние.
Неуравновешенность живых систем с окружающей средой поддерживается непрерывным потоком извне.
Неравновесное состояние живых систем поддерживается за счет извлечения ими из окружающей среды отрицательной энтропии. Назначение обмена живых систем с окружающей средой - освободиться от производимой положительной энтропии и извлечь отрицательную, т.е. повысить упорядоченность системы.
В экосистемах работают два различных механизма, производящих упорядоченность: статический, создающий порядок из беспорядка, и механизм, создающий порядок из порядка более низкого уровня.
Для растений характерен первый механизм выработки негоэнтропии, при котором за счет солнечной энергии в хлорофилле происходит повышение упорядоченности деградировавших веществ - фотосинтез.
Высшие животные и человек питаются высоко упорядоченными органическими соединениями (хотя энергии до высшего трофического уровня доходит мало, т.к. на каждом промежуточном уровне теряется до 90 %, но она поступает в концентрированной форме). Здесь работает второй механизм.
Жизнь - это единственный на Земле естественный самопроизвольный процесс, в котором энтропия уменьшается - за счет затрат даровой солнечной энергии. Необходимо только понимать, что энтропия уменьшается в конкретной локальной зоне, а в окружающей среде она из-за этого возрастает, т.е. осуществляется рост упорядоченности в других ее частях.
КПД преобразования солнечной энергии в энергию химических связей органических соединений очень мал. Но энергия солнца дается даром и главное, что растения и все живое обладают механизмом концентрирования избыточной энергии.
Необходимо подчеркнуть, что для поддержания внутренней упорядоченности в системе необходим не только постоянно действующий внешний источник энергии, но и наличие хорошо развитых диссипативных (рассеивающих) структур у самой системы.
Низкая энтропия в живых системах достигается постоянным и эффективным рассеиванием легко используемой концентрированной энергии. Упоминавшееся ранее нами клеточное дыхание высокоупорядоченной биомассы может рассматриваться в качестве основной диссипативной структуры экосистем.
Итак, экосистемы и организмы представляют собой открытые неравновесные термодинамические системы, постоянно обменивающиеся с окружающей средой энергией и веществом, уменьшая этим энтропию внутри себя, но увеличивая энтропию вовне.
Для описания "поведения" энергии в экосистемах нельзя употреблять термин круговорот энергии, а следует говорить о потоке энергии, поскольку в отличие от циклического движения веществ превращения энергии идут в одном направлении. Энергия, однажды использованная каким-либо образом, превращается в тепло и утрачивается для экосистемы.
Живые замкнутые термодинамические системы невозможны.
Каждый живой компонент, будь то организм или экосистема, должен получать от своей среды на входе постоянный приток энергии.
Энергия и продуктивность экосистем
Мы рассмотрели поток энергии в экосистеме качественно. Рассмотрим теперь процесс переноса энергии количественно с помощью его модели. Эта модель универсальна, т.е. применима к любой живой структуре, будь то растение, животное, микроорганизм, популяция или трофическая группа какого-либо уровня.
Пусть есть живая структура. Она имеет биомассу (массу органического вещества). Любое количество органического вещества эквивалентно некоторому количеству энергии (эту энергию можно извлечь, разрушив химические связи органического вещества). Например, 1 г. сухого вещества позвоночного животного (мясо) заключает 23.5 кДж.
Общее поступление энергии в систему (источник энергии) обозначен буквой I. Для автотрофов это солнечное излучение, для гетеротрофов - органическая пища, для некоторых видов живого требуется и то и другое, причем в определенных пропорциях.
Не вся энергия I, поступающая в биомассу В, подвергается превращению. Часть ее может пройти через биологическую структуру, не включаясь в метаболизм (обмен веществ), например, часть энергии света проходит через растительность не усваиваясь. Это так называемая неиспользованная энергия, обозначенная НЭ. Используемая или ассимилированная биологической структурой энергия обозначена буквой А.
А = I - НЭ
Ключевая особенность рассматриваемой модели - разделение ассимилированной энергии А на компоненты R и Р. Та часть энергии, которая идет на окисление органики, поддержание жизнедеятельности биологической структуры и в конечном итоге теряется в виде тепла, называется клеточным дыханием R, а та часть, которая превращается в новое органическое вещество, называется продукцией Р. Компонент Р - это энергия, доступная следующему трофическому уровню.
Соединенные между собой, такие графические модели могут отразить пищевые цепи в обществе или энергетику биосистемы в целом, где каждый такой блок описывает определенный трофический уровень в пищевой цепи.
Из рассмотренной модели видно, что в процессе жизнедеятельности сообщества создается и расходуется органическое вещество, т.е. каждая экосистема обладает определенной продуктивностью.
Продуктивность - это скорость образования органического вещества (биомассы). Различают несколько видов продуктивности.
Валовая первичная продуктивность системы определяется как скорость, с которой лучистая энергия Солнца усваивается организмами - продуцентами, в основном зелеными растениями в процессе фотосинтеза.
Однако в процессе жизнедеятельности растений часть созданного органического вещества расходуется на дыхание и, следовательно, накапливается меньше биомассы, чем ее было создано в результате фотосинтеза. Накопленное экологической системой вещество за вычетом того, которое израсходовано на дыхание, составляет чистую первичную продуктивность сообщества.
Органическое вещество, образующее чистую первичную продуктивность, доступно гетеротрофам. Консументы тоже создают органическое вещество (это выражается в частности в их росте). Они создают его за счет чистой первичной продуктивности и питания консументами более низкого уровня. Скорость накопления органического вещества на уровнях консументов называется вторичной продуктивностью.
В результате последовательности превращений энергии в пищевых цепях каждое сообщество приобретает определенную трофическую структуру. Трофическая структура сообщества отражает соотношение между продуцентами, консументами (отдельно первого, второго и т.д. порядков) и редуцентами, выраженное или количеством особей живых организмов, их биомассой, или заключенной в них энергией.
Графически трофическую структуру сообщества представляют обычно в виде пирамиды. Основанием пирамиды служит первый трофический уровень - уровень продуцентов, а последующие уровни образуют соответствующие этажи пирамиды (высота всех блоков - этажей одинакова, а длина пропорциональна числу, биомассе или энергии на соответствующем уровне).
Реальные пищевые цепи в естественных условиях могут быть очень длинными и ветвистыми.
Пирамида энергий является наилучшим графическим изображением структуры экосистемы, поскольку она отражает динамику системы, т.е. скорость прохождения энергии через пищевые цепи.
Пирамиды чисел и биомасс отражают статику системы, т.е. характеризуют количество или биомассу организмов в определенный промежуток времени.
Химический состав живого вещества и круговорот биогенов в экосистемах
Давно установлено, что растительный и животный мир состоят из тех же химических элементов, что и воздух, вода, минералы горных пород и почв. Наиболее важны среди них углерод - (С), водород - (Н), кислород - (О), азот - (N), фосфор - (Р) и сера - (S).
Значительную роль в построении живого вещества имеют калий (К), магний (Mg), кальций (Са), а также микроэлементы: Fe, Mn, B, Zn, Cu, Mo, Cl и др.
При сравнении живого и неживого очевидно, что, хотя слагающие их элементы одинаковы, характер организации атомов в молекулах существенно различается. В воздухе, воде и минералах они образуют относительно простые неорганические соединения, в живых организмах сложные органические (белки, углеводы, жиры, липиды и нуклеиновые кислоты, например ДНК* (дезоксирибонуклеиновая кислота), представляющие собой генетический материал организма).
Фундаментальные биологические процессы роста и размножения продуцентов представляют собой на химическом уровне поглощение из окружающей среды углерода, водорода, кислорода и других элементов в виде простых неорганических соединений и построение из этих атомов сложных систем органических молекул, т.е. тканей тела. Консументы осуществляют аналогичные химические преобразования, но начиная с более сложных веществ, получаемых с пищей.
Разложение и горение - это процессы, обратные росту, ведущие к разрушению сложных органических молекул и построению из этих атомов простых неорганических. Например, при сжигании сахара атомы углерода, водорода и кислорода, из которых он состоит, образует двуокись углерода и воду. Поскольку для этого требуется дополнительный кислород, говорят о процессе окисления.
При описании роста, разложения горения мы говорим лишь о перегруппировке атомов, а не об их изменении. Это свойственно всем химическим реакциям. Универсальный характер такого наблюдения позволяет говорить об одном их законов природы, а именно законе сохранения массы. Сейчас он формулируется следующим образом: атомы в химических реакциях никогда не исчезают, не образуются и не превращаются друг в друга; они только перегруппировываются с образованием различных молекул и соединений. (Атомы могут изменяться при высокоэнергетических ядерных реакциях, что и лежит в основе ядерной энергетики. Однако при биологических и химических процессах такие реакции исключены, поэтому с учетом вышесказанной оговорки закон сохранения массы можно считать универсальным).
Круговорот биогенов в экосистемах
Следствием закона сохранения вещества следует считать круговорот биогенов в экосистемах.
Мы уже отмечали, как четко взаимодействуют друг с другом продуценты, консументы, детритофаги и редуценты, поглощая и выделяя различные вещества. Органика и кислород, образуемые фотосинтезирующими растениями, - это как раз то, что нужно консументам для питания и дыхания. А выделяемые консументами углекислый газ и минеральные вещества мочи - как раз те биогены, которые необходимы растениям-продуцентам.
Мы видим, что в экосистеме получение ресурсов и избавление от отходов происходят в рамках круговорота всех элементов.
Этот принцип гармонирует с законом сохранения массы. Поскольку атомы не исчезают, не возникают и не превращаются один в другой, они могут использоваться бесконечно в самых различных соединениях, и запас их никогда не истощится. Именно это и происходит в естественных экосистемах. Рассмотрим такой круговорот более детально на примере трех важнейших элементов - углерода, фосфора и азота.
Круговорот углерода
Круговорот углерода (углекислоты) - процесс освобождения и связывания двуокиси углерода (СО2), включая ее растворение в воде океанов, идущий практически по двум циклам - океаническому и на суше, объединение между которыми происходит через атмосферную углекислоту. Баланс углерода в биосфере положителен в связи с антропогенными выбросами. Хотя в последнее время наблюдается некоторое замедление роста концентрации СО2 еще на 12-20%.
В ходе фотосинтеза атомы углерода включаются в состав глюкозы и других органических веществ, из которых построены все растительные ткани. В дальнейшем они переносятся по пищевым цепям и образуют ткани всех остальных живых существ экосистемы. Однако шансы отдельно взятого атома углерода "побывать" в течение одного цикла в составе многих организмов крайне малы, так как при каждом переходе с одного трофического уровня на другой велика вероятность, что содержащая его органическая молекула будет расщеплена в процессе клеточного дыхания для получения энергии. При этом атомы углерода вновь поступают в окружающую среду в составе углекислого газа, завершив один цикл и, приготовившись начать следующий. Аналогичным образом атомы углерода возвращаются в атмосферу в результате сжигания органики.
Еще одна важная и интересная особенность круговорота углерода состоит в том, что в
далекие геологические эпохи (сотни миллионов лет назад) значительная часть фотосинтезирующего органического вещества не использовалась ни консументами, ни редуцентами, накапливалась и постепенно погребалась под различными минеральными осадками. Находясь в земле миллионы лет, этот детрит под действием высоких температуры и давления превращался в нефть, природный газ и уголь; во что именно - зависело от исходного материала, продолжительности и условий пребывания в земле. Теперь мы в огромных количествах добываем это ископаемое топливо для обеспечения энергетических потребностей нашего индустриального общества, а, сжигая его, в определенном смысле завершаем круговорот углерода. Но одновременно резко возрастает концентрация углекислого газа в воздухе: его поступление туда существенно превышает поглощающие возможности современных растений. Это обстоятельство чревато серьезными климатическими последствиями.
Круговорот фосфора
Этот элемент входит в состав генов и молекул, переносящих энергию внутри клеток. В различных минералах фосфор содержится в виде неорганического фосфатиона (РО4)-3. Фосфаты растворимы в воде, но не летучи. Растения поглощают (РО4)-3 из водного раствора и включают в состав различных органических соединений, где фосфор выступает в форме так называемого органического фосфата. По пищевым цепям он переходит от растений ко всем прочим организмам экосистемы. Но опять же - при каждом переходе велика вероятность окисления содержащего фосфор соединения в процессе клеточного дыхания для получения организмом энергии. Когда это происходит, фосфат в составе мочи или ее аналога вновь поступает в окружающую среду, после чего снова может поглощаться растениями и начинать новый цикл.
В случае фосфора газовой фазы и, следовательно, "свободного возврата" в экосистему нет. Попадая в водоемы, фосфат насыщает, а иногда и перенасыщает их экосистемы. Обратного пути, по сути дела, нет. Конечно, что-то может снова вернуться на сушу с пометом рыбоядных птиц, но это очень незначительная часть общего количества, оказывающаяся к тому же вблизи побережья. Иногда океанические отложения фосфата со временем поднимаются над поверхностью воды в результате геологических процессов, но это требует миллионов лет.
Следовательно, фосфат и другие минеральные биогены почвы циркулируют в экосистеме лишь в том случае, если содержащие их "отходы" жизнедеятельности откладываются в местах поглощения данного элемента. В естественных экосистемах так по большей части и происходит. Однако мы создали нашу собственную экосистему человека, в которой урожай с извлеченными из почвы биогенами перевозится на большие расстояния к потребителям. А человеческие "отходы", содержащие эти питательные вещества, сбрасываются в водоемы.
Круговорот азота
Круговорот азота несколько сложнее, поскольку включает как газовую, так и минеральную фазу. Основная часть атомов азота находится в воздухе, который на 78 % состоит из одноименного газа (N2). Однако растения не могут усваивать его непосредственно; для этого азот должен входить в состав ионов аммония (NH4)+1 или нитрата (NO3)-1. К счастью, некоторые бактерии и ряд синезеленых водорослей (цианобактерий) способны превращать газообразный азот в аммонийную форму в ходе так называемой азотфиксации. Важнейшую роль среди азотфиксирующих организмов играют бактерии, живущие в клубеньках на корнях бобовых растений. По пищевым цепям органический азот, т.е. входящий в состав органических молекул, передается от бобовых растений другим организмам экосистемы.
Когда в процессе клеточного дыхания белки и другие содержащие азот органические соединения расщепляются, азот выделяется в среду главным образом в аммонийной (NH4)+1 форме. Некоторые бактерии могут переводить ее в нитратную форму (NO3)-1. Важно то, что, обе эти формы могут усваиваться любыми растениями. В результате азот совершает круговорот как минеральный биоген.
Однако такая минерализация обратима, поскольку другие почвенные бактерии постепенно превращают нитраты снова в газообразный азот (N2). Правда, часть его окисляется в воздухе - во время грозовых разрядов и поступает в почву с дождевой водой, но таким способом его фиксируется в 10 раз меньше, чем с помощью бактерий.
Значит, все естественные экосистемы зависят от азотфиксирующих организмов, поэтому крайне важна роль симбиотических бактерий в клубеньках бобовых растений. Это семейство включает огромное число представителей - от клевера (обычно на лугах и в степях) до тропических деревьев и пустынных кустарников. В каждой крупной наземной экосистеме - от дождевых экваториальных лесов до тундры есть характерные для нее виды бобовых. Интересно отметить, что бобовые обычно первыми заселяют гари. Без них процесс рекультивации будет идти значительно медленнее из-за недостатка в почве доступного азота. В водных экосистемах круговорот азота выглядит сходным образом, но здесь в роли основных азотфиксаторов выступают сине-зеленые водоросли.
Люди научились создавать искусственные экосистемы, выращивая урожаи кукурузы, пшеницы и других зерновых культур без участия бобовых. При этом азот воздуха фиксируется на специальных химических заводах. Искусственно полученные аммоний и нитрат представляют собой основные ингредиенты минеральных удобрений. Однако их высокая цена вынуждает многих фермеров реконструировать естественные условия, чередуя в севообороте бобовые и остальные культуры.
Большой и малый круговороты веществ
Солнечная энергия на Земле вызывает два круговорота веществ: большой или геологический, наиболее ярко проявляющийся в круговороте воды и циркуляции атмосферы; и малый или биологический. Оба круговорота взаимосвязаны и представляют единый процесс. Биологический круговорот веществ, втягивая в свои многочисленные орбиты костную среду, обеспечивает воспроизводство живого вещества и оказывает активное влияние на облик биосферы. В.И. Вернадский считал, что на земной поверхности нет химической силы, более постоянно действующей, а потому более могущественной по своим конечным последствиям, чем живые организмы, взятые в целом.
Основные принципы функционирования экосистем
Анализируя рассмотренные процессы функционирования экоситем, можно выделить его основные принципы и рассмотреть, как жизнедеятельность человека противоречит им.
Первый принцип. В естественных экологических системах использование ресурсов и избавление от отходов осуществляется в рамках круговорота всех элементов.
Второй принцип. Экосистемы существуют за счет не загрязняющей среду и практически вечной солнечной энергии, количество которой относительно постоянно и избыточно.
Третий принцип. На конце длинных пищевых цепей не может быть большой биомассы.
Четвертый принцип. Стабильность экосистемы определяется числом связей между видами в трофической сети, т.е. чем разнообразней видовой состав, тем устойчивее экосистема.
Экологические факторы
Неживая и живая природа, окружающая растения, животных и человека, носит название среды обитания. Она включает много отдельных компонентов, влияющих на организмы. Эти элементы среды называются экологическими факторами или факторами воздействия. Реальное значение разных факторов в жизни организмов не одинаково. Некоторые из них особенно важны и незаменимы. Их принято называть условиями существования.
По природе происхождения выделяют абиотические, биотические и антропогенные факторы.
Абиотические факторы - это свойства неживой природы, которые прямо или косвенно влияют на живые организмы.
Биотические факторы - это все формы воздействия живых организмов друг на друга. Каждый организм испытывает прямое или косвенное влияние других организмов, вступает в связь с представителями своего или других видов - растениями, животными, микроорганизмами, зависит от них самих и оказывает на них воздействие.
Раньше к биотическим факторам относили и воздействие человека на организмы, однако в настоящее время выделяют особую категорию факторов, порождаемых человеком.
Антропогенные факторы - это все формы деятельности человеческого общества, которые приводят к изменению природы как среды обитания, других видов и непосредственно сказываются на их жизни.
Таким образом, каждый живой организм испытывает влияние неживой природы, организмов других видов, в том числе и человека, и в свою очередь оказывает воздействие на каждую из этих составляющих. Действие всех факторов в совокупности, во взаимосвязи формирует и поддерживает устойчивость экосистемы; нарушение хотя бы одного из них может явиться причиной ее разрушения.
Экологические факторы действуют на организмы разными путями. В простейшем случае имеет место прямое влияние. Например, солнечные лучи освещают неподвижно лежащую ящерицу и тело ее согревается. С другой стороны, очень часто экологические факторы влияют косвенно или опосредованно, причем порой через многие промежуточные звенья. Например, сочетание высокой температуры воздуха с низкой влажностью и отсутствием дождей приводит к засухе. В результате на обширных пространствах выгорает растительность и травоядные дикие животные, например, антилопы, которые способны выносить жару и сухость, лишаются пищи и вынуждены либо мигрировать, либо погибать от голода и жажды. Это обстоятельство, в свою очередь, Сказывается на крупных хищных зверях и на трупоядных животных.
Закономерности воздействия экологических факторов
Несмотря на многообразие экологических факторов и различную природу их происхождения существуют некоторые общие правила и закономерности из воздействия на живые организмы. Рассмотрим эти универсальные правила.
Принцип лимитирующих факторов
Для жизни организмов необходимо определенное сочетание условий. Если все условия среды обитания благоприятны, за исключением одного, то именно это условие становится решающим для жизни рассматриваемого организма. Оно ограничивает (лимитирует) развитие организма, поэтому называется лимитирующим фактором.
Первоначально было установлено, что развитие живых организмов ограничивает недостаток какого-либо компонента, например, минеральных солей, влаги, света и т.п. В середине XIX в. немецкий химик-органик Либих первым экспериментально доказал, что рост растения зависит от того элемента питания, который присутствует в относительно минимальном количестве. Он назвал это явление законом минимума.
В современной формулировке закон минимума звучит так: выносливость организма определяется самым слабым звеном в цепи его экологических потребностей.
Закон толерантности
Однако, как выяснилось позже, лимитирующим может быт не только недостаток, но и избыток фактора (гибель урожая из-за дождей, перенасыщение почвы удобрениями и т.п.). Понятие о том, что наравне с минимумом лимитирующим фактором может быть и максимум, ввел спустя 70 лет после Либиха американский зоолог Шелфорд, сформулировавший закон толерантности. Согласно этому закону лимитирующим фактором процветания организма (вида) может быть как минимум, так и максимум экологического воздействия, а диапазон между ними определяет величину выносливости (предел толерантности) или экологическую валентность организма к данному фактору. Принцип лимитирующих факторов справедлив для всех типов живых организмов - растений, животных, микроорганизмов и относится не только к абиотическим, но и к биотическим факторам. Так, лимитирующим фактором для развития организмов данного вида может стать конкуренция со стороны другого вида. В земледелии, например, лимитирующим фактором часто становятся вредители, сорняки, а для некоторых растений лимитирующим фактором развития является недостаток (или отсутствие) представителей другого вида, например, опылителей (в Калифорнию из Средиземноморья завезли новый вид инжира, но он не плодоносил, пока оттуда же не завезли единственный для него вид пчел - опылителей).
Воздействие экологического лимитирующего фактора
Благоприятный диапазон воздействия экологического фактора называется зоной оптимума (нормальной жизнедеятельности). Чем значительнее отклонение от оптимума, тем больше данный фактор угнетает жизнедеятельность организма (зона угнетения). Максимально и минимально переносимые значения фактора - это критические точки, за пределами которых существование организма уже невозможно.
Представители разных видов сильно отличаются друг от друга, как по положению оптимума, так и по экологической валентности по отношению к разным экологическим факторам.
Для большинства видов оптимум биологической активности ограничен жесткими рамками. Сохранение должного уровня биологической активности обеспечивается гомеостатическими механизмами на уровне особи или популяции.
Широкую экологическую валентность вида по отношению к абиотическим факторам среды обозначают добавлением к названию фактора приставки "эври", например, животные, способные выносить значительные колебания температуры, называются эвритермными. Неспособность переносить значительные колебания факторов или узкая экологическая валентность характеризуется приставкой "стено", например, стенотермные животные.
Виды, для существования которых необходимы строго определенные экологические условия, называют стенобиотными, а приспосабливающихся к различной экологической обстановке - эврибиотными.
По особенностям реакции на изменение условий среды выделяют виды-конформисты и виды-регуляторы. У видов-конформистов в ответ на смену экологической ситуации происходят изменения определенных структурных и физиологических параметров (посткилотермные животные - пример). Виды-регуляторы противостоят колебаниям среды, поддерживая постоянство своей собственной среды (гомостотермные животные). В природе нет абсолютных конформистов и регуляторов. Так, например, у человека реакции конформистского типа на изменение условий окружающей среды - увеличение частоты дыхания и сердечных сокращений при восхождении на высокую гору. Пребывание в горах в течение нескольких дней включает другой, долгосрочный гомеостатический механизм - увеличение числа эритроцитов.
Предел толерантности (экологическая валентность) организма часто изменяется при переходе от одной стадии развития к другой. Часто молодые организмы оказываются более уязвимыми и более требовательными к условиям среды, чем взрослые особи. Наиболее критическим с точки зрения воздействия различных факторов является период размножения: В этот период многие факторы среды часто становятся лимитирующими. Экологическая валентность для размножающихся особей, семян, эмбрионов, яиц, личинок и т.п. обычно уже, чем для взрослых не размножающихся растений или животных того же вида. Так, например, многие морские животные могут переносить солоноватую воду или пресную воду с высоким содержанием хлоридов, поэтому они часто заходят в реки вверх по течению. Но их личинки не могут жить в таких водах, так, что вид не может размножаться в реке и не обосновывается здесь постоянно. Многие птицы летят выводить птенцов в места с более теплым климатом и т.п.
Взаимодействие экологических факторов
Оптимальная зона и пределы выносливости организмов по отношению к какому либо фактору среды могут смещаться в зависимости от того в каком сочетании действуют одновременно другие факторы. Эта закономерность получила название взаимодействия факторов. Например, известно, что жару легче переносить при сухом, а не влажном воздухе, угроза замерзания значительно выше при морозе с сильным ветром, чем в безветренную погоду. Для роста растений необходим, в частности, такой микроэлемент как цинк, именно он часто оказывается лимитирующим фактором. Но для растений, растущих в тени, потребность в нем меньше, чем находящиеся на солнце, т.е. происходит компенсация действия факторов.
Однако эта взаимная компенсация имеет определенные пределы и полностью заменить один из факторов другим нельзя. Полное отсутствие воды или хотя бы одного из основных элементов минерального питания делает жизнь растений невозможной, несмотря на самые благоприятные сочетания других условий. Отсюда следует вывод, что все условия среды, необходимые для поддержания жизни, играют главную роль и любой фактор может ограничивать возможности существования организмов (закон равнозначности всех условий жизни).
Необходимо отметить, что каждый фактор неодинаково влияет на разные функции организма. Условия, оптимальные для одних процессов (например, роста) могут являться зоной угнетения для других (например, размножение) и выходить за пределы толерантности (т.е. приводить к гибели) для третьих (например, гибель садово-огородных культур в весенние заморозки при их цветении). Поэтому жизненный цикл в соответствии, с которым организм в определенные периоды осуществляет преимущественно те или иные функции (питание, рост, размножение, расселение и т.п.) всегда согласован с сезонными изменениями факторов среды обитания.
Среди большого количества законов, определяющих взаимодействие отдельного индивида или особи с окружающей его средой выделим правило соответствия условий среды генетической предопределенности организма. Оно утверждает, что вид организмов может существовать до тех пор и постольку, поскольку окружающая его природная среда соответствует генетическим возможностям приспособления этого вида к ее колебаниям и изменениям.
Каждый вид живого возник в определенной среде, в той или иной степени приспособился к ней и дальнейшее его существование возможно лишь в ней или близкой к ней среде. Резкое и быстрое изменение среды жизни может привести к тому, что генетические возможности вида окажутся недостаточными для приспособления к новым условиям (на этом основана одна из гипотез вымирания крупных пресмыкающихся) с резким изменением абиотических условий на планете: крупные организмы менее изменчивы, чем мелкие, поэтому для адаптации им нужно гораздо больше времени. В связи с этим коренные преобразования природы опасны для ныне существующих видов, в том числе для самого человека.
Биотические факторы
Жизнедеятельность организмов определяется не только условиями внешней среды, которые будут рассмотрены ниже, но и отношениями с другими организмами. Непосредственное живое окружение организма составляет его биотическую среду, а факторы этой среды называются биотическими. Представители каждого вида способны существовать в таком окружении, где связи с другими организмами обеспечивают им нормальные условия жизни.
Формы биотических отношений
Выделяют различные формы биотических отношений. Если обозначить положительные результаты отношений для организма знаком "+", отрицательные результаты знаком "-", а отсутствие результатов - "0", то встречающиеся в природе типы взаимоотношений между живыми организмами можно представить в виде таблицы 1.
Таблица 1. Формы биотических отношений
Тип взаимодействия
Результат взаимодействия для вида
Общий характер взаимодействия
А
В
1. Конкуренция
-
-
Взаимное подавление обоих видов
2. Хищничество
+
-
Взаимодействие благоприятно для популяции А и неблагоприятно для В. (Особи хищников А крупнее, чем особи жертвы В)
3. Паразитизм
+
-
Взаимодействие благоприятно для популяции А и неблагоприятно для В. (особи паразита А мельче, чем особи хозяина В)
4. Аменсализм
-
0
Популяция В подавляет популяцию А, но сама не получает пользы от воздействия
5. Коменсализм
+
0
Популяция А (коменсам) получает пользу от объединения, популяции В это объединение безразлично
6. Симбиоз
+
+
Взаимодействие, благоприятное для А и В
7. Нейтрализм
0
0
Ни одна популяция не влияет на другую
Эта схематичная классификация дает общее представление о разнообразии биотических отношений. Рассмотрим примеры отношений различных типов.
Конкуренция является в природе наиболее всеохватывающим отношением, при котором две популяции или две особи в борьбе за необходимые для жизни условия воздействуют друг на друга отрицательно.
Конкуренция может быть внутривидовой и межвидовой. Внутривидовая борьба происходит между особями одного и того же вида. Межвидовая конкуренция имеет место между особями разных видов.
Конкурентное взаимодействие может касаться пространства, пищи или биогенных элементов, света, используемых веществ, зависимости от хищников, подверженности болезням и многих других типов взаимодействия. Межвидовая конкуренция, вне зависимости от того, что лежит в ее основе, может привести к установлению равновесия между двумя видами, либо к замене популяции одного вида популяцией другого, либо к тому, что один вид вытеснит другой в иное место или же заставит его перейти на использование иной пищи.
Конкурентные отношения отчетливо наблюдаются не только на межвидовом, но и внутривидовом (или популяционном) уровне. При росте популяции, когда численность ее особей приближается к насыщению, вступают в действие внутренние физиологические механизмы регуляции: возрастает их смертность, снижается плодовитость, возникают стрессовые ситуации, драки и т.п. Изучением этих вопросов занимается популяционная экология.
Хищничество и паразитизм наблюдается в случаях, когда отношения двух видов популяций благоприятны для одного из них и неблагоприятны для другого. Четко разграничить оба эти случая не всегда легко. Хищником можно назвать животное или растение, которое уничтожает жертвы, как правило, меньших размеров, чем оно само. Паразит - это организм, который питается за счет хозяина, более крупного, чем он, к которому он прикрепляется и причиняет ему вред.
Хищничество связано с активным поиском и энергичными способами овладевания сопротивляющейся добычей (жертвой). Для этого типа взаимоотношений характерно наличие разнообразных экологических адаптаций как у жертвы (окраска, панцири, шипы и т.д.), так и у хищников (развитие органов чувств, быстрота реакции, скорость передвижения и др.). Взаимодействие между хищником и жертвой влияет на их плотность и видовой состав, таким образом, осуществляется биологическая регуляция популяций.
Отметим, что когда два вида связаны отношениями паразитизма или хищничества, разрушительный эффект оказывается, в общем, небольшим в случае, когда популяции развивались вместе в стабильной в течение длительного времени среде. Однако действие окажется совершенно разрушительным, приводящим к полному исчезновению хозяина или жертвы, если два вида стали контактировать недавно или резко изменилась среда. Например, распахивая и возделывая новые территории и загрязняя их, человек оказался главной причиной исчезновения многих видов.
От паразитизма отличается аменсализм, при котором один вид причиняет вред другому, не извлекая при этом для себя никакой пользы. Чаще всего это те случаи, когда причиняемый вред заключается в изменении среды. Человек, разрушая и загрязняя окружающую среду, превращает большинство видов в аменсалов.
Симбиоз - длительное, неразделимое и взаимовыгодное отношение двух или более видов организмов. Например, жвачные животные (коровы, олени) переваривают клетчатку с помощью бактерий. Стоит, только, удалить этих симбионтов и животные погибнут от голода.
Другим видом положительных взаимодействий между двумя видами является комменсализм - извлекая из хозяина значительную пользу (пища, убежище), виды (комменсалы), не приносят ему никакой выгоды или заметного вреда. Например, многочисленные виды насекомых встречаются исключительно в муравейниках, норках грызунов, гнездах птиц, используя их как местообитание с более благоприятным микроклиматом.
Нейтрализм - ни одна из популяций не оказывает на другую никакого влияния: никак не сказывается на росте его популяций, находящихся в равновесии, и на их плотность. В действительности бывает довольно трудно при помощи наблюдений и экспериментов в природных условиях убедиться, что два вида абсолютно независимы один от другого.
Закономерности проявления биотических взаимодействий
Обобщая проявления биотических отношений в экосистемах можно выделить их основные закономерности:
1) отношение между живыми организмами является одним из основных регуляторов численности и пространственного распределения организмов в природе;
2) негативные взаимодействия между организмами в большей мере проявляются на начальных стадиях развития сообщества или в нарушенных природных условиях, в недавно сформировавшихся или новых ассоциациях вероятность возникновения сильных отрицательных взаимодействий больше, чем в старых ассоциациях;
3) установлено, что два одинаковых в экологическом отношении и потребностях вида сосуществовать в одном месте не могут и рано или поздно один конкурент вытесняет другого. Это так называемый принцип исключения или принцип Гаузе. (Если же близкородственные виды живут в одном месте, то они используют разные ресурсы или активны в разное время и т.п.).
Из принципа Гаузе следует, что каждый вид в природе занимает определенное своеобразное место. Оно определяется положением вида в пространстве, выполняемыми им функциями в сообществе и его отношениям к абиотическим условиям существования. Это место, занимаемое видом или организмом в экосистеме, называется экологической нишей. Образно говоря, если место обитания - это как бы адрес организмов данного вида, то экологическая ниша - это профессия, функциональная роль вида в месте его прописки;
4) в естественных условиях ни один вид не стремится (и не может привести) к уничтожению другого. Более того, исчезновение какого-либо естественного "врага" из экологической системы может привести к вымиранию того вида, за счет которого развивается этот "враг".
Классификация абиотических факторов
К абиотическим факторам среды относятся климатические, эдафические, топографические, гидрофизические и гидрохимические факторы.
Из климатических факторов основное экологическое значение имеют температура, влажность (количество осадков) и солнечная радиация. Вторичные климатические факторы (ветер, атмосферное давление, прозрачность атмосферы и др.) играют меньшую роль.
Факторы водной среды - физикохимические свойства воды, кислотность среды, содержание минеральных и органических веществ, а также кислорода, течение и т.п.
Почвенные (эдафические) факторы - механический и органический состав, минеральные вещества, кислотность, влажность, газовый состав почвы и т.п.
Факторы рельефа (топография) - высота над уровнем моря, крутизна и ориентация склонов, перепады высот.
Атмосферно-климатические факторы
Температура - наиболее важный фактор
Температура является наиболее важным климатическим фактором. От нее зависит интенсивность обмена веществ организмов и их географическое распространение. Любой организм способен жить в пределах определенного интервала температур. И хотя для разных видов организмов эти интервалы различны (эвритермные и стенотермные организмы), для большинства из них амплитуда оптимальных температур, при которых жизненные функции осуществляются наиболее активно, сравнительно небольшая.
Диапазон температур, в которых может существовать жизнь, составляет примерно 3000 (от -2000С до +1000С). Но большинство видов и большая часть активности приурочены к еще более узкому диапазону температур. Определенные организмы, особенно в стадии покоя, могут существовать, по крайней мере, некоторое время, при очень низких температурах, отдельные виды микроорганизмов, главным образом бактерии и водоросли, способны жить и размножаться при температурах близких к точке кипения. Верхний предел для бактерий горячих источников - 880С, для синезеленых водорослей - 800С, а для самых устойчивых рыб и насекомых - 500С. Как правило, верхние предельные значения фактора оказываются более критичными, чем нижние, хотя многие организмы вблизи верхних пределов диапазона толерантности функционируют более эффективно.
У водных животных диапазон толерантности к температуре обычно более узок по сравнению с наземными животными, так как диапазон колебаний температуры в воде меньше, чем на суше.
С точки зрения воздействия на живые организмы крайне важен характер изменчивости температуры. Температура, колеблющаяся от 100С до 200С (в среднем 150С), не обязательно действует на организм также как постоянная температура 150С. Жизнедеятельность организмов, которые в природе обычно подвергаются воздействию переменных температур, подавляется полностью или частично или замедляется под действием постоянной температуры. Это подтверждают эксперименты Шелфорда (1929) и Паркера (1930), обнаруживших, что яйца, личинки и куколки яблочной плодожорки в условиях колеблющейся температуры развиваются на 7-8 % быстрее, чем при постоянной. Стимулирующий эффект переменных температур, по крайней мере в умеренной зоне можно считать установленным.
Таким образом, температура является важным тоже время лимитирующим фактором. Температурные ритмы в значительной степени контролируют сезонную и суточную активность растений и животных. Температура часто создает зональность и стратификацию в водных и наземных местообитаниях.
Вода и влажность
Вода физиологически необходима для любой протоплазмы. С экологической точки зрения она служит лимитирующим фактором как в наземных местообитаниях, так и в водных.
Все живые организмы, в зависимости от потребности их в воде, а, следовательно, и по различиям местообитания, подразделяются на ряд экологических групп: 1) водные или гидрофильные - живут постоянно в воде; 2) гигрофильные - живут в очень влажных местообитаниях; 3) мезофильные - отличаются умеренной потребностью в воде и 4) ксерофильные - живут в сухих местообитаниях.
Количество осадков и влажность - основные величины, измеряемые при изучении этого фактора.
Количество осадков зависит, в основном, от путей и характера перемещения воздушных масс. Простой пример: Ветры, дующие с океана, составляют большую часть своей влаги на обращенных к океану склонах, в результате чего за горами создается "дождевая тень", способствующая формированию пустыни.
Распределение осадков по времени года - крайне важный лимитирующий фактор для организмов. Условия, создающиеся в результате равномерного распределения осадков, совершенно иные, чем при распределении осадков во время одного сезона. В этом случае растениям и животным приходится переносить периоды длительной засухи. Как правило, неравномерное распределение осадков по временам года встречается в тропиках и субтропиках, где нередко выражены влажный и сухой сезоны. В тропическом поясе сезонный ритм влажности регулирует сезонную активность организмов аналогичного сезонному ритму тепла и света в условиях умеренного пояса. Роса может представлять собой значительный, а в местах с малым выпадением дождей и очень важный вклад в общее количество осадков.
Влажность - параметр, характеризующий содержание водяного пара в воздухе. Абсолютной влажностью называют количество водяного пара в воздухе, выраженное через массу воды на единицу массы воздуха. В связи с зависимостью количества пара, удерживаемого воздухом, от температуры давления введено понятие относительной влажности, т.е. отношение содержащегося в воздухе пара к насыщаемому пару при данных температуре и давлении. Так как в природе существует суточный ритм влажности (повышение ночью и снижение днем), а также ее колебания по вертикали и горизонтали, этот фактор, наряду со светом и температурой, играет важную роль в регулировании активности организмов. Влажность изменяет эффекты воздействия температуры.
Например, при условиях влажности, близких к критическим, температура оказывает более выраженное лимитирующее влияние. Аналогично, влажность играет более критическую роль, если температура близка к предельным значениям. Крупные водоемы значительно смягчают климат суши, так как для воды характерна большая скрытая теплота парообразования и таяния. Фактически существуют два основных типа климата: 1) континентальный, с крайними значениями температуры и влажности, и 2) морской, которому свойственны менее резкие колебания, что объясняется смягчающим влиянием крупных водоемов.
Доступный запас поверхностной воды зависит от количества осадков в данном районе, но эти величины не всегда совпадают. Так, пользуясь подземными источниками, куда вода поступает из других районов, животные и растения могут получать больше воды, чем ее поступает с осадками. И наоборот, дождевая вода иногда сразу же становится недоступной для организмов.
Солнечное излучение
Солнечное излучение совершенно необходимо живой природе, т.к. является единственным внешним источником энергии. Надо иметь в виду, что спектр электромагнитного излучения Солнца весьма широк и его частотные диапазоны различным образом воздействуют на живое вещество. Интегральная плотность излучения Солнца на среднем расстоянии до Земли (а.е.=150*106км) равна 0.14 Вт/см2, при этом средняя температура солнечного диска равна 58000K.
Спектр распределения энергии излучения Солнца за пределами земной атмосферы показывает, что около половины солнечной энергии излучается в ИК-области, 40 % - в видимой (от 0.4 до 0.7 мкм) и 10 % - в ультра- и рентгеновской областях.
Земная атмосфера, включая озоновый слой, селективно, т.е. избирательно по частотным диапазонам, поглощает энергию электромагнитного излучения Солнца и до поверхности Земли доходит в основном излучение с длиной волны от 0.3 до 3 мкм. Более длинно- и коротковолновое излучение поглощается атмосферой. С увеличением зенитного расстояния Солнца возрастает относительное содержание ИК-излучения (от 50 до 72 %).
Для живого вещества очень важны качественные показатели излучения (длина волны, интенсивность и продолжительность воздействия). Интенсивность фотосинтеза несколько варьируется с изменением длины волны света.
Например, при прохождении света через воду красная и синяя части спектра отфильтровывается и получающийся зеленоватый свет слабо поглощается хлорофиллом. Однако красные водоросли имеют дополнительные пигменты (фитоэритриты), позволяющие им использовать эту энергию и жить на большой глубине, чем смогли бы зеленые водоросли.
Интенсивность света, падающего на автотрофный ярус, управляет всей экосистемой, влияя на первичную продуктивность. И у наземных, и у водных растений фотосинтез связан с интенсивностью света линейной зависимостью до оптимального уровня светового насыщения, за которым во многих случаях следует снижение интенсивности фотосинтеза. Отдельные растения и целые сообщества приспосабливаются к различным интенсивностям света, становясь "адаптированными к тени" (диатомовые, фитопланктон) или "адаптированными к прямому солнечному свету".
Продолжительность светового дня или фотопериод является "реле времени" или пусковым механизмом, включающим последовательность физиологических процессов, приводящих к росту и цветению многих растений, линьке и накоплению жира, миграции и размножению у птиц и млекопитающих и к наступлению диапаузы у насекомых.
Газовый состав атмосферы
В настоящее время атмосфера Земли имеет следующий состав: О2 - 21 %, N2 - 78 %, CO2 - 0.03 %, 0.97 % - инертные газы и примеси. Интересно, что концентрации О2 и СО2 являются до какой-то степени лимитирующими для животных видов и многих высших растений.
У ряда растений удается повысить эффективность фотосинтеза, повысив концентрации СО2. Жизнь млекопитающих и человека становится невозможной при концентрации О2 < 16 %.
Азот - важнейший биогенный элемент, участвующий в образовании белковых структур организмов. Озон (О3) является производной атмосферного кислорода и выполняет экранирующую роль по отношению к УФ части солнечного спектра, губительной для всего живого.
Скорость движения воздуха и барометрическое давление
В атмосфере никогда не бывает полного покоя. На суше лимитирующее воздействие на активность и даже на распространение организмов оказывает скорость ветра. Ветер способен изменять внешний вид растений особенно в тех местообитаниях, например в альпийских зонах, где лимитирующее воздействие оказывают другие факторы.
Уайкед (1958) экспериментально показал, что в открытых горных местообитаниях ветер лимитирует рост растений. Большое значение имеют бури, хотя их действие сугубо локально. Ураганы, да и обычные ветры, способны переносить животных и растения на большие расстояния и тем самым изменять состав сообществ.
Барометрическое давление, по-видимому, не является лимитирующим фактором непосредственного действия, однако оно имеет прямое отношение к погоде и климату, которые оказывают непосредственное лимитирующее воздействие.
Факторы водной среды
Водные условия создают своеобразную среду обитания организмов, отличающуюся от наземной прежде всего плотностью и вязкостью.
Плотность воды примерно в 800 раз, а вязкость примерно в 55 раз выше, чем у воздуха. Вместе с плотностью и вязкостью важнейшими физико-химическими свойствами водной среды являются: постоянные перемещения водных масс в пространстве, способствующие сохранению однородности физических и химических характеристик; температурная стратификация, т.е. изменение температуры по глубине водного объекта, режим, связанный периодическими изменениями температуры; прозрачность воды, определяющая световой режим под ее поверхностью.
От прозрачности зависит фотосинтез зеленых и пурпурных водорослей, фитопланктона, высших растений, а, следовательно, накопление органического вещества возможно лишь в пределах эвфотической зоны, где процессы фотосинтеза преобладают над процессами дыхания.
В водных местообитаниях количество кислорода, оксида углерода (IV) и других атмосферных газов, растворенных в воде и потому доступных организмам, сильно варьируется во времени. В водоемах с высоким содержанием органических веществ кислород является лимитирующим фактором первостепенной важности (СО2 = 1 %).
Запас кислорода в воде пополняется при помощи диффузии из воздуха и благодаря фотосинтезу водных растений. Содержание его меняется в воде в зависимости от времени суток, времени года и местоположения.
Небольшое увеличение концентрации СО2 в воде повышает интенсивность фотосинтеза и стимулирует процессы развития многих организмов. Высокие же концентрации СО2 являются лимитирующим фактором для животных, т.к. высокое содержание СО2 сопровождается низким содержанием О2. Например, рыбы весьма чувствительны к повышению концентрации СО2: при слишком высоком содержании свободного СО2 в воде многие рыбы погибают.
Концентрация водородных ионов (pН) тесно связана с карбонатной системой. Значение pН изменяется в диапазоне 0 < pH < 14; при pН = 7 среда нейтральная; pH < 7 - кислая, pH > 7 - щелочная. Если pH не приближается к крайним значениям, то сообщества способны компенсировать изменения этого фактора и толерантность сообщества к диапазону pH весьма значительна. pH может служить индикатором скорости общего метаболизма сообщества. В водах с низким pH содержится мало биогенных элементов, поэтому продуктивность здесь крайне мала.
Еще одним значимым абиотическим фактором в водных объектах является соленость (содержание карбонатов, сульфатов, хлоридов и т.д.). В пресных водах их мало, причем около 80 % составляют карбонаты. Содержание же минеральных веществ в Мировом океане составляет в среднем 35 г/л. Организмы открытого океана обычно стеногалинны, тогда как организмы прибрежных солоноватых вод в общем эвригалинны. Концентрация солей в жидкостях тела и тканях большинства морских организмов изотонична концентрации солей в морской воде, так что здесь не возникает проблем с осморегуляцией.
Течение не только сильно влияет на концентрацию газов и питательных веществ, но и прямо действует как лимитирующий фактор. Многие речные растения и животные морфологически и физиологически особым образом приспособлены к сохранению своего положения в потоке: у них есть вполне определенные пределы толерантности к фактору течения.
Гидростатическое давление в океане имеет большое значение. С погружением в воду на 10 м давление возрастает на 1 атм (около 105 Па). В самой глубокой части океана давление достигает 1000 атм. Многие животные способны переносить резкие колебания давления, особенно если в их теле нет свободного воздуха. В противном случае возможно развитие газовой эмболии. Высокие давления, характерные для больших глубин, как правило угнетают процессы жизнедеятельности.
Почвенные факторы
Почвой называют слой вещества, лежащий поверх горных пород земной коры. Русский ученый Докучаев в 1870 г. первым рассмотрел почву как динамическую, а не инертную среду. Он доказал, что почва постоянно изменяется и развивается, а в ее активной зоне идут физические, химические и биологически процессы. Почва формируется в результате сложного взаимодействия климата, растений, животных и микроорганизмов. Рост растений зависит от содержания необходимых веществ в почве и от ее структуры.
В состав почвы входят четыре основных структурных компонента: минеральная основа (обычно 50-60 % общего состава почвы), органическое вещество (до 10 %), воздух (15-25 %) и вода (25-35 %).
Минеральный скелет почвы - это неорганический компонент, который образовался из материнской породы в результате ее выветривания.
Свыше 50 % минерального состава почвы занимает кремнезем (кварц SiO2), 1-25 % приходится на глинозем (Al2O3), 1-10 % - на оксиды железа (Fe2O3), 0.1-5 % - на оксиды магния, калия, фосфора, кальция. Минеральные фрагменты, образующие вещество почвенного скелета, различны по размерам - песчинки диаметром 0.02-2 мм, ил (0.002-0.02 мм) и частицы глины (менее 0.002 мм). Их соотношение определяет структуру почвы. Глины и суглинки (содержат примерно равные количества глины и песка) обычно пригодны для роста растений, так как содержат достаточно питательных веществ и способны удерживать влагу. Песчаные почвы быстрее дренируются и теряют питательные вещества из-за выщелачивания (вымывания водой).
Органическое вещество почвы образует при разложении мертвых организмов, их частей (например, опавших листьев) и экскрементов. Не полностью разложившиеся органические остатки называются подстилкой, а конечный продукт разложения - аморфное вещество, в котором уже невозможно распознать первоначальный материал, - называется гумусом. Благодаря своим химическим и физическим свойствам гумус улучшает структуру почвы и ее аэрацию (воздухообмен), а также повышает способность удерживать воду и питательные вещества.
Почвенный воздух также, как и почвенная вода, находится в порах между частицами почвы. Порозность (объем пор) возрастает в ряду от глин к суглинкам и пескам. Обычно в воздухе почвы из-за дыхания населяющих ее организмов несколько меньше кислорода и больше диоксида углерода, чем в атмосферном воздухе. Если идет процесс заболачивания, то почвенный воздух вытесняется водой и условия становятся анаэробными. Почва постепенно становится кислой, так как анаэробные организмы продолжают вырабатывать СО2. Почва, если она небогата основаниями, может стать чрезвычайно кислой, а это наряду с истощением запасов кислорода неблагоприятно воздействует на почвенные микроорганизмы. Длительные анаэробные условия ведут к отмиранию растений.
Почвенные частицы удерживают вокруг себя некоторое количество воды. Часть ее, называемая гравитационной водой, может свободно просачиваться вниз сквозь почву. Это ведет к вымыванию из почвы различных минеральных веществ, в том числе азота. Вода может также удерживаться вокруг отдельных коллоидных (размером менее 1 мкм = 0,001 мм) частиц в виде тонкой прочной связанной пленки. Эту воду называют гигроскопической. Она адсорбируется на поверхности частиц за счет водородных связей. Эта вода наименее доступна для корней растений, и именно она последней удерживается в очень сухих почвах. Количество гигроскопической воды зависит от содержания в почве коллоидных частиц, поэтому в глинистых почвах их намного больше (примерно 15 % веса почвы), чем в песчанистых (примерно 0,5 %). По мере того, как накапливаются слои воды вокруг почвенных частиц, она начинает заполнять сначала узкие поры между этими частицами, а затем распространяется во все более широкие поры. Гигроскопическая вода постепенно переходит в капиллярную, которая удерживается вокруг почвенных частиц силами поверхностного натяжения. Капиллярная вода может подниматься по узким порам и канальцам от уровня грунтовых вод. Растения легко поглощают капиллярную воду, которая играет наибольшую роль в регулярном снабжении их водой. В отличие от гигроскопической эта вода легко испаряется. Тонкоструктурные почвы удерживают больше капиллярной воды, чем грубоструктурные.
Вода поступает в живые клетки почвенных организмов путем осмоса (односторонняя диффузия вещества через полупроницаемую мембрану, разделяющую два раствора различной концентрации).
Вода важна как растворитель питательных веществ и газов, поглощаемых из водного раствора корнями растений. Она принимает участие в процессе почвообразования.
Химические свойства почвы зависят от содержания минеральных веществ, которые находятся в ней в виде растворенных ионов. Концентрация в почве ионов водорода (рН) в среднем близка к нейтральному значению (рН = 7). Флора таких почв особенно богата видами. Известковые и засоленные почвы имеют рН = 8 - 9, а торфяные, кислые - до 4. На этих почвах развивается специфическая растительность.
В почве обитает множество видов растительных и животных организмов, влияющих на ее физико-химические характеристики: бактерии, водоросли, грибы или простейшие одноклеточные, черви и членистоногие.
Основные биохимические процессы протекают в верхнем слое почвы толщиной до 0,4 м, так как в нем обитает наибольшее количество микроорганизмов. Одни бактерии участвуют в цикле превращения только одного элемента, другие - в циклах превращения многих элементов. Если бактерии минерализуют органическое вещество (разлагают органическое вещество на неорганические соединения), то простейшие уничтожают избыточное количество бактерий. Дождевые черви, личинки жуков, клещи разрыхляют почву и этим способствуют ее аэрации. Кроме того, они перерабатывают трудно расщепляемые органические вещества.
Топографические факторы
Главным топографическим фактором является высота. С высотой снижаются средние температуры, увеличивается суточный перепад температур, возрастает количество осадков, скорость ветра и интенсивность радиации, понижаются атмосферное давление и концентрации газов. Все эти факторы влияют на растения и животных. В результате обычным явлением стала вертикальная зональность.
Горные цепи могут служить климатическими барьерами. Горы служат также барьерами для распространения и миграции организмов и могут играть важную роль изолирующего фактора в процессах видообразования.
Еще один важный топографический фактор - экспозиция склона. В северном полушарии склоны, обращенные на юг, получают больше солнечного света, поэтому интенсивность света и температура здесь выше, чем на дне долин и на склонах северной экспозиции. В южном полушарии имеет место обратная ситуация.
И, наконец, важным фактором рельефа является крутизна склона. Для крутых склонов характерны быстрый дренаж и смывание почв, поэтому здесь маломощные и более сухие. Если уклон превышает 350, почва и растительность обычно не образуются, а создаются осыпи из рыхлого материала.
Огонь как абиотический фактор периодического воздействия
В настоящее время экологи пришли к однозначному мнению, огонь (пожар) надо рассматривать как один из естественных абиотических факторов (может возникать, например, от молнии) наряду с температурой, атмосферными осадками, почвой и т.п.
Пожары как экологический фактор бывают различных типов и оставляют после себя различные последствия. Верховые или "дикие" пожары (т.е. очень интенсивные и не поддающиеся сдерживанию) разрушают всю растительность, да всю органику почвы, последствия же низовых пожаров совершенно иные. Верховые пожары оказывают лимитирующее действие на большинство организмов - биотическому сообществу приходится начинать все сначала, с того немногого, что осталось, и должно пройти много лет, пока участок снова станет продуктивным. Низовые пожары, напротив, обладают избирательным действием: для одних организмов они оказываются более лимитирующими, для других - менее и, таким образом, способствуют развитию организмов с высокой толерантностью к пожарам. Кроме того, небольшие низовые пожары дополняют действие бактерий, разлагая отмершие растения и ускоряя превращение минеральных элементов питания в форму, пригодную для использования новыми поколениями растений.
Если низовые пожары случаются регулярно раз в несколько лет, валежника на земле мало, это снижает вероятность возгорания крон. В лесах, не горевших более 60 лет, накапливается столько горючей подстилки, отмершей древесины, что при ее воспламенении верховой пожар почти неизбежен.
Растения выработали специальные адаптации к пожару также, как они сделали по отношению к другим абиотическим факторам. В частности, почки злаков и сосен скрыты от огня в глубине пучков листьев или хвоинок. В периодически выгорающих местообитаниях эти виды растений получают преимущества и огонь способствует их сохранению, избирательно благоприятствует их процветанию.
Широколиственные же породы лишены защитных приспособлений от огня, он для них губителен.
Таким образом, пожары поддерживают устойчивость лишь некоторых экосистем.
Реймерс Н.Ф. Надежды на выживание человечества: Концептуальная экология, М., Экология, 1992.
3.9. Экосистемные законы
Провести четкую грань между биоценозом, всегда занимающим какой-то определенный биотоп, и экосистемой, представляющей единство биоценоза и биотопа, довольно трудно. Распределение экологических обобщений, касающихся этих двух классов образований, по "ячейкам" параграфов весьма условно. Помимо известных концепций экосистемы А. Тенсли (1935) и биогеоценоза В.И. Сукачева (1940) была выдвинута точка зрения или, скорее, сформулировано правило Ф. Эванса (1956), предложившего использовать термин "экосистема" абсолютно "безразмерно" для обозначения любой надорганизменной живой системы, взаимодействующей с окружением. С точки зрения общей теории систем такой подход вполне логичен, однако семантически многие авторы термину "экосистема" придали значение именно биогеоценоза, т.е. элементарной экосистемы, и одновременно более высоких по иерархии надбиогеоценотических образований вплоть до экосистемы биосферы. Подразделения по иерархии более низкие для группового наименования не получили общепринятого обобщающего термина. Видимо, их можно называть биосистемами, что, вероятно, лучше, чем обозначать их термином организм (надорганизм), фактически рекомендованным в "Биологическом энциклопедическом словаре" (см. главу 1). Однако такое обозначение использовать непросто - в главе 2 биосистемами названы образования от организма до популяции, экобиосистемами - от особи до экологической пирамиды внутри существенной части биоценоза - его парцеллы, а образования биоценотического ряда вообще пока не поддаются обобщению в виде одного понятия, если не принять термин биоценосистемы.
Выделить законы функционирования только для уровня элементарной экосистемы - биогеоценоза - едва ли возможно. Поэтому граница между разделом 3.9. и следующим, посвященным закономерностям, характерным для биосферы, условна и проходит главным образом по такой характеристике биосферы как глобальность и большая условная замкнутость (при термодинамической открытости). Другие же экосистемы Земли вещественно почти не замкнуты, что налагает на действующие в них закономерности ряд ограничений.
3.9.1. Структура и функционирование экосистем
- Принцип экологической комплетарности (дополнительности)
- Принцип экологической конгруэнтности (соответствия)
- Принцип (закон) формирования экосистемы, или связи биотоп - биоценоз
- Закон однонаправленности потока энергии
- Закон внутреннего динамического равновесия
- Закон экологической корреляции
- Правило оптимальной компонентной дополнительности
- Принцип экологической (рабочей) надежности
- Принцип видового обеднения
Сложение экосистем в значительной мере зависит от их функциональной "предназначенности" и наоборот. Это банальное замечание сделано для того, чтобы настроить на восприятие принципа экологической комплентарности (дополнительности): никакая функциональная часть экосистемы (экологический компонент, элемент и т.п.) не может существовать без других функционально дополняющих частей. Происходит это в силу того, что каждая подсистема связана с другими согласно общему принципу дополнительности и группе закономерностей сложения систем (см. закон необходимого разнообразия, закон полноты составляющих, правило конструктивной эмерджентности и др.) Например, любой организм поглощает (ассимилирует) из внешней среды одни вещества и выделяет в нее продукты своей жизнедеятельности (диссимиляции). Если бы не было дополняющих видов, использующих продукты диссимиляции и восстанавливающих их до продуктов ассимиляции, через какой-то промежуток времени необходимые ресурсы жизни для организма были бы исчерпаны. Сходный принцип наблюдается и в эволюции любых систем (Правило системно-динамической комплементарности).
На основе эмпирических данных можно сформулировать близкий к предыдущему и расширяющий его принцип экологической конгруэнтности (соответствия): функционально дополняя друг друга, живые составляющие экосистемы вырабатывают для этого соответствующие приспособления, скоординированные с условиями абиотической среды, в значительной мере преобразуемой теми же организмами (биоклимат и т.п.). То есть средой их обитания - внешней и создаваемой ценозом. Действительно, скажем, виды, составляющие экосистемы пустыни, с одной стороны приспособлены к ее климатическим и другим абиотическим условиям, а с другой и к друг к другу. То же характерно для организмов любого биома и другого более низко или высоко стоящего в иерархии систем подразделения биосферы.
Ю.Н. Куражковский формулирует четвертый закон жизни: "постоянное существование организмов в любом ограниченном пространстве возможно только в экологических системах, внутри которых отходы жизнедеятельности одних видов организмов утилизируются другими видами". Несколько видоизменив смысл этого обобщения, предлагается принять формулировку очень близкого принципа (закона) формирования экосистемы (функционально-пространственной экологической целостности, связи биотоп - биоценоз): длительное существование организмов возможно лишь в рамках экологических систем, где их компоненты и элементы дополняют друг друга и соответственно приспособлены друг к другу. Это обеспечивает воспроизводство среды обитаний каждого вида и относительно неизменное существование всех экологических компонентов. Совершенно очевидно, что принцип формирования экосистемы есть суммарное отражение принципа экологической комплентарности (дополнительности) и принципа экологической конгруэнтности (соответствия). Очевидна его связь со всей группой правил сохранения среды обитания - видовой, на уровне биоценоза (относительной внутренней непротиворечивости) и биосферы.
При обсуждении закона (принципа) "энергетической проводимости" уже упоминался второй экологический закон, по Ю.Н. Куражковскому, "закон сохранения жизни: жизнь может существовать только в процессе движения через живое тело потока вещества, энергии и информации. Прекращение движения в этом потоке прекращает жизнь". Скорее, это общий диалектический постулат или, уже, действительно закон жизни, а не только экологии, хотя он справедлив и для любых экологических образований и вообще многих природных систем, даже непосредственно не связанных с живым.
Более специфичен для экологических и биоценотических систем закон однонаправленности потока энергии: энергия, получаемая сообществом (экосистемой) и усваиваемая продуцентами, рассеивается или вместе с их биомассой передается консументам первого, второго и т.д. порядков, а затем редуцентам с падением потока на каждом трофическом уровне в результате процессов, сопровождающих дыхание. Поскольку в обратный поток (от редуцентов к продуцентам) поступает ничтожное количество изначально вовлеченной энергии (не более 0,25, максимум 0,35%), говорить о "круговороте энергии нельзя. Существует лишь круговорот веществ, поддерживаемый потоком энергии.
Из закона однонаправленности потока энергии, конечно, с учетом ее рассеивания, а потому "бокового" использования в соседних экосистемах, и того обстоятельства, что энергия приходит в экосистему не только непосредственно от Солнца, но и опосредованно через вещественные абиотические экологические компоненты (атмосферу, воду, субстраты), вытекают многочисленные следствия и прежде всего правило (принцип) экологического дублирования, а также практически все другие обобщения, сведенные в упомянутом разделе. Однонаправленность потока энергии формирует в экосистемах относительно замкнутый круговорот веществ. Закономерности распространения информации также связаны с потоком энергии и перемещением вещества, но они пока еще мало изучены. Видимо, здесь большую роль играют горизонтальные связи между экосистемами, хотя информационные сети явно гуще и лучше налажены в пределах образований каждого уровня их иерархии. Пока еще поле обобщений в области экоинформатики почти не возделано. Тут непочатый край работы для специалистов.
Эмпирически более ясен вопрос о взаимоотношении энергии, вещества и информации внутри экосистем и отношении этого взаимодействия к их динамическим качествам. В начале 70-х гг. автором был сформулирован закон внутреннего динамического равновесия, а затем четыре основных следствия из него. Формулировка закона: вещество, энергия, информация и динамические качества отдельных природных систем (в том числе экосистем) и их иерархии взаимосвязаны настолько, что любое изменение одного из этих показателей вызывает сопутствующие функционально-структурные количественные и качественные перемены, сохраняющие общую сумму вещественно-энергетических, информационных и динамических качеств систем, где эти изменения происходят, или в их иерархии.
Действие закона внутреннего динамического равновесия совершенно четко связано с законом однонаправленности потока энергии. Именно ограниченность этого потока и специфические свойства формируют всю массу связей в экосистеме в их разнообразии. Поэтому и соблюдается экологический аналог законов сохранения массы и энергии.
Справедливость закона внутреннего динамического равновесия доказывается практикой ведения хозяйства и особенно характером региональных экологических катастроф типа сахельской, приаральской, азовской, кара-богазской, бассейнов Белого и Баренцева морей, волжско-каспийской и других. Этот закон - одна из основных путеводных нитей в управлении природопользованием, впрочем, нитей, долго игнорировавшихся.
Важные для практики следствия из закона внутреннего динамического равновесия приведем в том же порядке, что в словаре-справочнике "Природопользование":
1. Любое изменение среды (вещества, энергии, информации, динамических качеств экосистем) неизбежно приводит к развитию природных цепных реакций, идущих в сторону нейтрализации произведенного изменения или формирования новых природных систем, образование которых при значительных изменениях среды может принять необратимый характер.
2. Взаимодействие вещественно-энергетических экологических компонентов (энергия, газы, жидкости, субстраты, организмы-продуценты, консументы и редуценты), информации и динамических качеств природных систем количественно нелинейно, т.е. слабое воздействие или изменение одного из показателей может вызвать сильные отклонения в других (и во всей системе в целом).
Например, малое отклонение в составе газов атмосферы, ее загрязнение окислами серы и азота вызывают огромные изменения в экосистемах суши и водной среды. Именно оно привело к возникновению кислотных осадков, а с ними к деградации и гибели лесов в Европе и Северной Америке, обезрыбливанию озер Скандинавии, нарушению циклов развития морских организмов, личиночные стадии которых проходят на мелководьях. Столь же абсолютно незначительное изменение концентрации углекислого газа ведет к возникновению тепличного эффекта, а комплекс малых перемен формирует массовые размножения организмов.
3. Производимые в крупных экосистемах изменения относительно необратимы - проходя по их иерархии снизу вверх, от места воздействия до биосферы в целом, они меняют глобальные процессы и тем самым переводят их на новый эволюционный уровень (согласно закону необратимости эволюции Л. Долло).
4. Любое местное преобразование природы вызывает в глобальной совокупности биосферы и в ее крупнейших подразделениях ответные реакции, приводящие к относительной неизменности эколого-экономического потенциала (правило "тришкина кафтана"), увеличение которого возможно лишь путем значительного возрастания энергетических вложений (согласно закону снижения энергетической эффективности природопользования). Искусственный рост эколого-экономического потенциала ограничен термодинамической (тепловой) устойчивостью природных систем (закон одного процента).
Пока изменения среды слабы и произведены на относительно небольшой площади, они или ограничиваются конкретным местом, или "гаснут" в цепи иерархии экосистем. Но как только перемены достигают существенных значений для крупных экосистем, например, в ограниченных масштабах больших речных бассейнов или в размерах, ограниченных правилами одного и десяти процентов, они приводят к существенным сдвигам в этих обширных природных образованиях, а через них согласно 2-му следствию из обсуждаемого закона, и во всей биосфере Земли. Будучи относительно необратимыми, изменения в природе в конечном итоге оказываются и трудно нейтрализуемыми с социально-экономической точки зрения: их выправление требует слишком больших материальных средств и физических усилий. Иногда возникает даже ситуация "чем больше пустынь мы превратим в цветущие сады, тем больше цветущих садов мы превратим в пустыни". При этом в силу нелинейности процессов опустынивание по темпам значительно опережает создание "цветущих садов" и теоретически не может не опережать, поскольку такое создание базируется на нарушении компонентного равновесия в экосистемах.
Сдвигая динамически равновесное (квазистационарное) состояние природных систем с помощью значительных вложений энергии (например, путем распашки и других агротехнических приемов), люди нарушают соотношение экологических компонентов, достигая увеличения полезной продукции (урожая) или состояния среды, благоприятного для жизни и деятельности человека. Если эти сдвиги "гаснут" в иерархии природных систем (от элементарного до биосферы и экосферы планеты в целом) и не вызывают термодинамического разлада, положение благоприятно, или, во всяком случае, терпимо. Однако излишнее вложение энергии и возникающий в результате вещественно-энергетический разлад ведут к снижению природно-ресурсного потенциала вплоть до опустынивания территории, происходящего без компенсации: вместо цветущих садов возникают пустыни.
В связи с нелинейностью, неполной пропорциональностью взаимоотношения компонентов и возникновением цепных природных реакций ожидаемый при преобразовании природы эффект может не возникнуть или оказаться многократно более сильным, чем желательно. В первом случае местная реакция как бы начнет "скользить" по иерархии природных систем, "растворяться" в ней и, достигнув уровня всей биосферы или ее крупных подразделений, "исчезнет" (она становится неизмеряемой при наших возможностях распознавания перемен в природе). Во втором случае, наоборот, надсистемы усиливают процесс "сверху вниз", он делается острее, заметнее. Это заставляет при проектировании природопользовательских акций рассматривать не только местные вещественно-энергетические балансы, но учитывать вероятные изменения в надсистемах. В противном случае игнорирование обсуждаемого закона приводит к ошибкам в природопользовании.
Прямым следствием закона внутреннего динамического равновесия является более узкий по смыслу закон экологической корреляции: в экосистеме, как и в любом другом целостном природно-системном образовании с участием живого, все входящие в нее виды живого и абиотические экологические компоненты функционально соответствуют друг другу. Выпадение одного элемента системы, например, исчезновение вида, неминуемо ведет к исключению всех тесно связанных с этим элементом системы других ее частей и функциональному изменению целого в рамках внутреннего динамического равновесия. При этом выпадение элемента может вести и к включению нового блока, т.е. исключению с положительным, а не отрицательным знаком - внедрению в экосистему нового вида или их группы всегда вместе с их консорбентами. Действие закона экологической корреляции ведет к скачкообразным изменениям экологической устойчивости: при достижении порога изменения функциональной целостности происходит часто неожиданный срыв - экосистема теряет свойство надежности. Например, многократное увеличение концентрации загрязняющего вещества может не приводить к заметным последствиям, но затем ничтожная его прибавка приведет к катастрофе. В данном случае возникает экологический вариант закона "все или ничего" Х. Боулича. Загрязнитель, воздействуя на наименее устойчивые организмы и меняя цепь природных реакций, вызывает срыв в экологической корреляции, и экосистема начинает разрушаться как бы изнутри, т.е. саморазрушаться при минимальном изменении силы внешнего фактора.
Следствием закона экологической корреляции является правило оптимальной компонентной дополнительности: Никакая экосистема не может самостоятельно существовать при искусственно созданном значительном и перманентном избытке или недостатке одного из экологических компонентов. "Нормой" экологического компонента следует считать ту, которая обеспечивает экологическое равновесие определенного типа, позволяющее функционировать именно той экосистеме, которая эволюционно сложилась и соответствовать балансу в природной надсистеме и всей иерархии природных систем на данной единице пространства (в конкретном биотопе).
Длительное изменение количества и качества одного из экологических компонентов ведет к замене одной экосистемы другой, при этом не всегда хозяйственно желательной. Как правило, природные изменения экологических компонентов бывают непродолжительными и не выходят за рамки нормы реакции экосистемы. Искусственные же изменения нередко бывают перманентными, отнюдь не всегда соотнесены с выносливостью экосистем и часто приводят к их деградации.
В экологии одним из самых запутанных вопросов является соотношение между сложностью (разнообразием видов) и устойчивостью (надежностью) биотических сообществ и экосистем. Предложены сотни моделей математической интерпретации этих зависимостей. Однако многие экологи обращают внимание на отсутствие прямых и постоянных связей между разнообразием и надежностью, включая устойчивость экосистем. Р. Риклефс указывает на то, что "экологи удивительно невежественны во всем, что касается устойчивости природных систем" и формулирует "один из основных экологических принципов: эффективность сообщества и его стабильность возрастают прямо пропорционально той степени, в которой составляющие его популяции в процессе эволюции приноровились друг к другу".
К этому заключению следует добавить, что в экосистемах соответствие необходимо не только между популяциями, но и экологическими компонентами, а также между данной экосистемой и ее положением в надсистеме, всей их иерархии. Попытки найти парные связи в одних случаях успешны, в других - нет, и это вполне естественно в сложных иерархически построенных совокупностях, какими являются экосистемы.
Исходя из данных, накопленных экологией, с учетом вышеприведенных обобщений возможно сформулировать принцип экологической (рабочей) надежности: эффективность экосистемы, ее способность к самовосстановлению и саморегуляции (в пределах естественных колебаний) зависит от ее положения в иерархии природных образований (краевое, маргинальное, положение невыгодно), степени взаимодействия ее компонентов и элементов (их экологической комплементарности, конгруэнтности, характера динамического равновесия, экологической корреляции и оптимальности в компонентной дополнительности), а также от частных приспособлений организмов (размеры, продолжительность жизни, скорость смены поколений, отношение продуктивности к биомассе и т.п.), составляющих биоту экосистемы. Разнообразие, сложность и другие морфологические черты экосистемы имеют неодинаковое значение и подчинены степени ее эволюционной и сукцессионной зрелости. Если снижение разнообразия приводит к резкому дисбалансу в "притертости" частей экосистемы, а это случается достаточно часто, то упрощение системы чревато заметным снижением ее надежности. Эти качественные изменения не могут уловить количественные модели. В связи с этим нет взаимопонимания между "классическими" экологами и эколого-математиками. Как правило, быстро развивающиеся (кратковременно-производные), сравнительно недавно нарушенные экосистемы оказываются надежнее климаксовых из-за повышенной устойчивости к внешним воздействиям (это свойство было ранее шутливо определено нами как "за битого двух небитых дают").
Сформулированное обобщение в свете имеющихся данных кажется очевидным. Однако привести лаконичный пример его реализации довольно трудно, можно апеллировать лишь к монографиям по экологии и знаниям читателей. Сам поиск прямых связей между сложностью экосистемы и ее надежностью кажется данью упрощенному мышлению технократического типа. "Простота" любой биосистемы лишь иллюзия. На самом деле мы имеем дело с квазипростыми образованиями с высокой степенью сложности. Элементарный редукционизм в этом случае недопустим. Можно говорить лишь об общих тенденциях, всегда учитывая специфику каждого из рассматриваемых образований.
Сформулированы следующие принципы видового обеднения:
1) консорционной целостности: с исчезновением из экосистемы вида, образующего консорцию, исчезают и многие консорбенты, не входящие в другие консорции той же или другой территориально и функционально близкой экосистемы (принцип "никто не гибнет в одиночку");
2) биологического замещения: вновь внедрившийся в экосистему вид всегда, вырабатывая свою экологическую нишу, сужает возможности менее конкурентоспособных видов и тем ведет к их исчезновению или лишь слегка видоизменяет экологические ниши функционально близких видов, создавая предпосылки для сокращения их численности или, наоборот, массового размножения ("принцип незваного гостя"); при этом меняется все сообщество, включая даже как будто не связанные с внедрившимся видом формы (в силу смещения общего экологического равновесия);
3) смены трофических цепей (сетей): исчезнувшая трофическая цепь (сеть) видов сменяется цепью (сетью) эколого-энергетических аналогов, что позволяет экосистеме усваивать и перерабатывать поступающую извне энергию. Этот принцип вытекает из правила экологического дублирования и кратко звучит: "экосистема не терпит пустоты";
4) неопределенности хозяйственного значения смены видов: при замене трофических цепей (сетей) вновь проникающие в экосистему (в редких случаях возникающие в ней) виды могут быть как желательными, так и нежелательными в хозяйственном и медицинском отношении (принцип "старый друг лучше новых двух").
Перечисленные принципы имеют большое практическое значение. Вместе с тем они как бы перебрасывают мостик от структурно-функциональных свойств экосистем к их динамике.
3.9.2. Динамика экосистем
- Принцип сукцессионного замещения
- Теория мозаичного климакса
- Закон последовательности прохождения фаз развития
- Закон сукцессионного замедления
- Правило максимума энергии поддержания зрелой экосистемы, или правило Г. Одума и Р. Пинкертона
- Принцип "нулевого максимума", или минимизации прироста в зрелой экосистеме
- Принцип "сукцессионного очищения", или стабилизации и минимизации видового состава климакса
- Правило увеличения замкнутости биогеохимического круговорота веществ в ходе сукцессии
- Правило сукцессионного мониторинга, или степени завершенности сукцессии
- Закон эволюционно-экологической необратимости
Сложение экосистемы - динамический процесс. Ее формируют виды, дополняющие друг друга и соответствующие другим видам и условиям среды - сами по себе и в складывающейся совокупности. При этом происходит выработка экологических ниш - функциональное размежевание в возникающем множестве или его дополнение при малом разнообразии. На жизнепригодном субстрате биотические сообщества последовательно формируют закономерный ряд экосистем, ведущий к наиболее устойчивой в данных условиях климаксовой в чисто природных условиях или узловой при природно-антропогенном режиме фазе. В этом смысл принципа сукцессионного замещения. При антропогенном или зоогенном поддержании какой-то из фаз сукцессии возникают параклимаксы. Спор о возможности поликлимаксов кажется чисто умозрительным. В строго определенных условиях возможен только моноклимакс. Если возникают различные варианты климакса - поликлимакс, значит, наблюдаются отклонения в среде обитания. Столь категорическое утверждение следует из парадигмы детерминированности развития. Оно всегда однонаправлено согласно значительному блоку общесистемных закономерностей.
Сказанное не означает, что регионально не существует мозаика климаксовых экосистем, обусловленная топографическими, геоморфологическими и другими условиями среды. Именно такая мозаика лежит в основе теории мозаичного климакса, развитой Р. Уиттекером в 1957 г. Картосхема биотопов, основанная на ландшафтно-геоботанической, а, вернее, экосистемной идеологии, может служить иллюстрацией к теории Р. Уиттекера: сукцессионные ряды в долине реки, в сосняках, протянувшихся вдоль нее, в лесах склонов гор и на их верхних частях составляют достаточно четкую мозаику. Если бы была приведена карта восстановленной растительности, эта закономерность выступила бы еще ярче. Но это не значит, что, скажем, климакс приречного ельника может как угодно варьировать.
Смена фаз сукцессии идет в соответствии с определенными правилами. Каждая фаза готовит среду для возникновения последующей. Тут действует закон последовательности прохождения фаз развития. Существует тенденция считать, что этот закон в рамках сукцессионного процесса не действует или не обязательно действует. При этом происходит смешение понятий естественного развития и смены экосистем и искусственной культивации, в основном леса. Нередко, оставив экосистему в рамках полуестественного развития и только посадив саженцы лесной породы, хозяйственник не получает ожидаемого эффекта - посадки гибнут. При этом причины могут быть самыми различными, но всегда они связаны с неподготовленностью желаемой фазы развития реальной предыдущей.
Когда экосистема приближается к состоянию климакса, в ней, как и во всех равновесных системах, происходит замедление всех процессов развития. Это положение сформулировано в виде закона сукцессионного замедления: процессы, идущие в зрелых равновесных экосистемах, находящихся в устойчивом состоянии, как правило, проявляют тенденцию к снижению темпов. При этом восстановительный тип сукцессий меняется на вековой их ход, т.е. саморазвитие идет в пределах климакса или узлового сообщества. Например, восстановительная сукцессия темнохвойного леса в южной тайге может идти следующим образом. Сначала быстро меняется ряд луго-кустарниковых фаз с образованием березового или осинового леса. Затем может произойти либо замещение этого леса сосной, либо прямое вытеснение елью (в горной южной Сибири пихтово-кедровым лесом). Сосняк может составить узловое сообщество, и процесс смены пород резко тормозится, но в других случаях сосну относительно быстро вытесняет ель или пихтово-кедровый лес. Эти климаксовые образования отнюдь не лишены тенденций к развитию, но оно резко заторможено. Вековая сукцессия идет по типу заболачивание - разболачивание или остепнение - расстепнение (по климатическим характеристикам, а не по доминирующему характеру растительности). Возможен и вариант образования вечной мерзлоты, и тогда темно-хвойный лес в Сибири и на Дальнем Востоке будет медленно вытеснен лиственничным. В речных долинах на Дальнем Востоке может образоваться параклимакс из чозении. В этом случае процессы развития также весьма замедлены. Совершенно очевидно, что при хозяйственных акциях учет скорости естественных процессов обязателен.
Эмпирический закон сукцессионного замедления есть следствие правила Г. Одума и Р. Пинкертона, или правила максимума энергии поддержании зрелой системы: сукцессия идет в направлении фундаментального сдвига потока энергии в сторону увеличения ее количества, направленного на поддержание системы. В свою очередь правило Г. Одума и Р. Пинкертона базируется на правиле максимума энергии в биологических системах, сформулированном А. Лоткой. Позже оно было расширено Р. Маргалефом. Вопрос этот хорошо разработан в фундаментальной монографии Ю. Одума "Основы экологии" как и доказательство принципа "нулевого максимума", или минимизации прироста в зрелой экосистеме: экосистема в сукцессионном развитии стремится к образованию наибольшей биомассы при наименьшей биологической продуктивности. Этот факт совершенно очевидно вытекает из правила Г. Одума и Р. Пинкертона и служит эмпирической его иллюстрацией. В общем правилом Г. Одума и Р. Пинкертона объясняется и принцип "сукцессионного очищения", или стабилизации и минимизации видового состава климакса: разнообразие стремится к пику на ранних или средних фазах сукцессии, а затем снижается в климаксе. Этот постулат Р. Маргалефа явно связан с увеличеним потока энергии, поддерживающего климакс. Совершенно очевидна его связь с принципами экологического высвобождения и экологической компрессии.
В ходе сукцессии происходит увеличение замкнутости биогеохимических круговоротов веществ. Хотя все биогеохимические круговороты не полностью замкнуты, но степень незамкнутости варьирует от очень больших величин (практически стремящихся к бесконечности на поверхности планеты, не защищенной растительностью) до значения 104. Примерно за 10 лет с момента начала восстановления растительного покрова разомкнутость круговоротов уменьшается со 100 до 10%, а далее она еще больше снижается, в климаксовой фазе достигая минимума. Правило увеличения замкнутости биогеохимического круговорота веществ в ходе сукцессии, как совершенно очевидно, нарушается антропогенной трансформацией растительности и вообще естественных экосистем. Это ведет к длинному ряду аномалий в биосфере и ее подразделениях.
Снижение разнообразия видов в климаксе не означает малой его значимости. В этом отношении вопрос Ю. Одума в вышеупомянутой работе: "представляет ли собой разнообразие только "приправу" к жизни или оно необходимо для долгой жизни всей экосистемы, в которую входят человек и природа" кажется не совсем корректным. Именно разнообразие видов формирует сукцессию, ее направление, обеспечивает заполненность реального пространства жизнью. Если бы оно было несущественным и имело только "эстетическое" значение, то виды, составляющие комплекс, не могли бы сформировать сукцессионный ряд, и постепенно, с разрушением климаксовых экосистем произошло бы полное опустынивание планеты. Ведь значение разнообразия функционально не только в статике, но и в динамике.
Там, где разнообразие видов недостаточно для формирования биосреды, служащей основой нормального, естественного хода сукцессионного процесса, а сама среда резко нарушена (по глубине или частоте нарушений), сукцессия не достигает фазы климакса, а заканчивается узловым сообществом, параклимаксом, длительно- или даже кратковременнопроизводным сообществом. Качество среды данной территории, суммарная напряженность антропогенного воздействия отражается на степени завершенности сукцессионного ряда. Чем глубже нарушенность среды какого-то пространства, тем на более ранних фазах оканчивается сукцессия. Это правило сукцессионного мониторинга (индикации состояния среды), или степени завершенности сукцессии, как кажется, имеет вполне практическое значение, особенно в картрировании экологического состояния территорий и акваторий.
Достижение климакса при потере одного или группы видов в результате их уничтожения (реже вымирания, тоже, как правило, связанного с антропогенным исчезновением местообитаний) не есть полное восстановление природной обстановки. Фактически это новая экосистема, так как в ней возникли новые связи и утеряны многие старые, сложилась иная "притертость" видов. Вернуться в старое состояние экосистема не может, так как утерянный вид невосстановим. Это позволяет сформулировать закон эволюционно-экологической необратимости: экосистема, потерявшая часть своих элементов или сменившаяся другой в результате дисбаланса экологических компонентов, не может вернуться к первоначальному своему состоянию в ходе сукцессии, если в ходе изменений произошли эволюционные перемены в экологических элементах (сохранившихся или временно утерянных) или один либо группа видов исчезла навсегда или на (системно) долгий срок. Если какие-то виды утеряны в промежуточных фазах сукцессии, то эта потеря может быть функционально скомпенсирована, но лишь частично. При снижении разнообразия за критический уровень ход сукцессии искажается и фактически климакс, идентичный прошлому, достигнут быть не может.
Закон эволюционно-экологической необратимости важен для оценки характера восстановленных экосистем. При потере элементов это, по сути дела, совершенно экологически новые природные образования с вновь образовавшимися закономерностями и связями. Например, перенос давно выбывшего из состава экосистемы вида в ходе его реакклиматизации - отнюдь не механическое его возвращение. Фактически это внедрение нового вида в обновленную экосистему.
Закон эволюционно-экологической необратимости подчеркивает фактическую направленность эволюции не только на уровне биосистем, но и на всех иных иерархических уровнях сложения биоты.
Вопросы для обсуждения или письменных заданий
Ниже приведены примерные вопросы, которые Вы можете задать учащимся, студентам. Эти вопросы дают им возможность высказать свое отношение к проблемам окружающей среды, а также помогают закрепить и просуммировать то, что они узнали из рекомендуемой литературы.
I. Полагаете ли Вы, что человеческие существа, являясь лишь одним среди миллионов других видов, населяющих экосистемы на Земле, имеют моральное право на их изменение в своих собственных интересах, обрекая тем самым другие виды на вымирание?
2. В свете принципов второго закона сохранения энергии скажите, оправданна ли с термодинамической и экологической точек зрения зависимость человека от мяса животных как источника протеина? Почему?
3. Как Вы полагаете, оправданно ли с позиций окружающей среды введение в экосистему организмов из других экосистем?
Можете ли Вы привести примеры, когда это было оправданно, даже если привнесенные виды нарушили целостность экосистем, например, когда исчезли один или более местных видов?
Практические задания
Эти задания дадут учащимся и студентам возможность применить материал раздела к реальным жизненным ситуациям.
I. Выберите район вблизи Вашего дома или учебного заведения. Определите тип биома, характерный для данного района. Подготовьте письменное описание биома и характерных для него видов, основанное на Ваших собственных наблюдениях и дополнительных исследованиях. Насколько богата экосистема выбранного района с точки зрения числа и разнообразия местных видов растений и животных? Оказала ли антропогенная деятельность заметное влияние на природную экосистему?
2. Сколько трофических уровней Вы можете обнаружить в рассмотренной выше экосистеме? Какие виды являются первичными продуцентами, какие первичными консументами? Имеются ли в экосистеме консументы второго, третьего или более высокого уровня?
3. Вычертите диаграммы биогеохимических циклов углерода и азота. Определите, каким образом в каждом цикле задействованы микроорганизмы.
Контрольные вопросы
1. Какие абиотические факторы связаны соотношением влажности и температуры?
а. Механические
б. Почвенные
в. Климатические В
2. В лесном массиве был создан заповедник. К какому экологическому фактору можно отнести этот факт?
а. Биотический
б. Абиотический
в. Антропогенный В
3. Найдите сумму факторов, определяющих жизнь:
а. Минеральные соли, рельеф
б. Температура, вода, свет
в. Влияние человека Б
4. Под биотическим фактором понимают:
а. Сумму воздействия живых организмов
б. Воздействие человека на организмы
в. Факторы неживой природы А
5. Линька птиц и перелеты в теплые страны связаны с:
а. Понижением температуры воздуха
б. Изменением влажности воздуха
в. Изменением продолжительности дня В
6. Какие приспособления способствуют выживанию животных в засушливых районах?
а. Анабиоз
б. Накопление жиров
в. Интенсивное образование метаболической воды в организме в результате окислительных реакций В
7. Береза бородавчатая относится к растениям:
а. Гелиотрофным (светлолюбивым)
б. Сциофитным (теневые)
в. Гелиосцитныи (теневыносливым) В
8. Свет для прорастания семян:
а. Совсем не нужен
б. Нужен гелиофитам
в. Нужен только паразитическим растениям Б
9. Найди из перечисленных факторов антропогенный фактор:
а. Свет
б. Взаимосвязи между особями в популяции
в. Развитие промышленности В
10. Наиболее важны биохимические адаптации к температуре дня:
а. У животных
б. У растений и микроорганизмов
в. У человека
г. Все перечисленное Б
11. Отмершие листья деревьев образуют опад, который служит местом обитания и пищей для многих организмов. К каким факторам относится это явление?
а. Антропогенные
б. Биотические
в. Абиотические
г. Комплексное действие факторов В
12. Из перечисленных факторов выберите абиотические факторы:
а. Влажность, свет, температура
б. Вырубка леса
в. Влияние организмов друг на друга А
13. Приспособления к дневному и ночному способу жизни у животных возникли в связи с изменением:
а. Освещенности
б. Температуры
в. Влажности А
14. Фотопериодизм - это реакция организма на изменение:
а. Температуры воздуха
б. Влажности воздуха
в. Соотношение дня и ночи В
15. Выберите приспособления растений к перезимовке:
а. Увеличение количества воды в тканях (для предотвращения обезвоживания)
б. Накопление углеводов в клетках (приводящее к связыванию воды)
в. Прекращение тока ассимиляторов Б
16. Из перечисленных животных выберите тех, которые имеют непостоянную температуру тела
а. Насекомые
б. Млекопитающие
в. Птицы А
17. К абиотическим факторам природы относятся:
а. Климат
б. Взаимосвязи между особями в популяции
в. Воздействие человека на организм А
18. Что является для растений и животных сигналом к сезонным изменениям?
а. Температура
б. Длина дня
в. Количество пищи Б
19. Антропогенный фактор среды, это:
а. Фактор неживой природы
б. Непосредственное взаимодействие человека на природу
в. Живые организмы, влияющие друг на друга Б
20. Какие из перечисленных факторов относятся к искусственному загрязнению:
а. Вулканические извержения
б. Выхлопные газы автомобилей, смог
в. Пыльные бури Б
21. Назовите организм, у которого физиологические процессы ускоряются в связи с повышением температуры окружающей среды:
а. Воробей
б. Кошка
в. Гусеница бабочки капустной белянки
г. Камбала В
22. Какой из перечисленных компонентов экосистемы является редуцентом?
а. Муха
б. Ящерица
в. Дождевой червь В
23. Указать, какая из перечисленных пищевых цепей составлена правильно?
а. Муха - лягушка - уж
б. Уж - лягушка - муха
в. Лягушка - муха - уж А
24. Численность белки в еловых лесах в первую очередь зависит от?
а. Количества хищников
б. Суровости зимы
в. Урожая семян В
25. Большую роль в биомассе играют организмы:
а. Животных
б. Растений
в. Микроорганизмов Б
26. От чего зависит степень конкуренции?
а. От ареала конкурентов
б. От количества конкурентов
в. От сильного сходства или различия потребностей конкурентов В
27. Что такое симбиоз?
а. Прямое уничтожение жертвы
б. Совместное существование, при котором каждый вид извлекает пользу от связей с другими видами
в. Форма взаимоотношений, при которых один вид использует другой в качестве источника питания, принося вред хозяину Б
28. Какое из перечисленных взаимодействий относится к хищничеству?
а. На теле хозяина живут вши
б. Волки поедают мелких травоядных
в. Насекомые питаются нектаром цветковых растений Б
29. Азотфиксирующие бактерии относятся к:
а. Продуцентам
б. Консументам
в. Редуцентам Б
30. Многие организмы создают среду для других организмов или меняют условия их обитания. Такие связи между организмами называются:
а. Трофические
б. Топические
в. Мутализм Б
31. Птицы сороки предупреждают копытных об опасности. О каком типе взаимодействия идет речь?
а. Мутуализм
б. Межвидовая взаимопомощь
в. Симбиоз Б
32. Выбери пример нейтрализма
а. Корраловые полипы и одноклеточные водоросли
б. Белки и насекомоядные птицы в одном лесу, в отсутствии угнетающих факторов
в. Змеи и амфибии Б
33. Выбери пример паразитизма:
а. Береза и гриб подберезовик
б. Повилика и растения
в. Лишайники Б
34. Выбери пример хищничества:
а. Змей и птичьи яйца
б. Корраловые полипы и одноклеточные водоросли
в. Минога и треска А
35. Выбери пример конкуренции:
а. Акула и рыбы прилипалы
б. Деревья и микоризные грибы
в. Ель в березовом лесу В
36. Роль хищников в природе заключается в том, что:
а. Уменьшение количества популяций-жертв
б. Поддержание состава и численности популяций-жертв на оптимальном уровне
в. Избирательное уничтожение животных с низкой способностью добывать себе корм
г. Все перечисленное Г
37. Многие хищники не способны предотвратить вспышки численности жертв. С чем это связано?
а. Хищники имеют меньший жизненный цикл
б. Во время вспышки численности "жертв" плодовитость хищников уменьшается
в. У хищников скорость роста популяции меньше, чем у жертв
г. Все перечисленное В
38. Светолюбивые травы, растущие под елью, страдают от сильного затенения, тогда как самому дереву это безразлично. Какой тип взаимодействия характеризует данный пример?
а. Протокооперация
б. Конкуренция
в. Аменсализм
г. Мутуализм Б
39. Выберите из приведенного ниже списка организм, относящийся к консументам:
а. Муравей
б. Водосбор лекарственный
в. Гадюка
г. Гриб-навозник В
40. Какой из данных вариантов может быть примером паразитизма?
а. Вши, клещи, грибы
б. Рак-отшельник и актиния
в. Березка под пологом елового леса А
41. Волки ежегодно убивают около 25% оленей. К какой форме взаимодействия относится это явление?
а. Конкуренция
б. Хищничество
в. Паразитизм Б
42. Борьба внутри вида или между особями различных видов за пищу, ареал обитания, это:
а. Конкуренция
б. Хищничество
в. Паразитизм А
43. Найдите правильный ответ: совместное существование, при котором каждый вид извлекает пользу от связи с другим видом?
а. Нейтрализм
б. Мутуализм
в. Конкуренция Б
44. За что не конкурируют земные организмы?
а. За пищу
б. За воздух
в. За территорию Б
45. Важное значение для хищников имеет:
а. Количество пищи
б. Качество пищи
в. Разнообразие пищи А
46. Какое из перечисленных взаимодействий относится к симбиозу?
а. Воробей поедает червей
б. Кукушка откладывает яйца в гнезда мелких воробьиных
в. В рубце одного из отделов сложного желудка жвачных обитает несколько килограммов бактерий В
47. Какие из приведенных ниже организмов могут выступать в роли и консументов и редуцентов?
а. Трутовик
б. Петров крест
в. Индюк А
48. Выберите условие существования конкуренции?
а. Ограничение общих ресурсов
б. Избыток общих ресурсов
в. Существует независимо от ресурсов А
49. Какой из ниже приведенных примеров относится к мутуализму?
а. Комар сосет кровь человека
б. Насекомые собирают нектар и опыляют растения
в. Воробей питается ягодами, способствуя распространению семян Б
50. Птицы кормятся насекомыми-паразитами на коже носорога, а их взлет служит носорогу сигналом опасности. Какой из типов симбиотических связей характеризует этот пример?
а. Комменсализм
б. Мутуализм
в. Протокооперация Б
51. Как называется форма взаимоотношений, при которой один вид получает какое-либо преимущество, выгоду при этом, не приносят другому ни вреда, ни пользы?
а. Мутуализм
б. Комменсализм
в. Нейтрализм Б
52. Выбери пример мутуализма:
а. Кедровая сосна и кедровка
б. Береза и гриб-трутовик
в. Гриб-фитофтора и картофель А
53. Выбери пример комменсализма:
а. Осина и микорида подосиновика
б. Мелкие рыбки в полости голотурий
в. Растения семейства росянковые и насекомые Б
54. В чем суть воздействия особей-конкурентов одного вида друг на друга?
а. В снижении плодовитости
б. В уменьшении скорости роста популяции
в. В приобретении новых качеств, способствующих выживанию
г. В полной гибели особей данного вида В
55. Из перечисленных признаков выбери признак, обязательный для типа взаимоотношений "хозяин-паразит"
а. Тесная связь с хозяином, как результат естественного отбора
б. Полное истощение и быстрая смерть хозяина
в. Нанесение вреда хозяину без пользы для себя
г. Извлечение пользы для себя без вреда хозяину А
56. Известно, что хищничество связано с активным поиском пищи, поэтому для него характерно наличие разнообразных экологических адаптаций. Из перечисленных, найдите адаптации, характерные для хищников:
а. Покровительственная окраска
б. Использование убежищ
в. Быстрота реакции
г. Наличие шипов, игл, панциря В
57. Крапива любит влажные места с плодородными почвами, теневынослива. Василек поселяется на различных почвах, в условиях хорошей освещенности. Какие на Ваш взгляд отношения могут быть между ними?
а. Симбиоз
б. Нейтрализм
в. Конкуренция
г. Паразитизм Б
58. Аллелопатия - это:
а. Химическое взаимовлияние растений друг на друга при помощи продуктов метаболизма
б. Взаимополезные связи двух видов, предполагающие обязательное сожительство
в. Потребление остатков пищи хозяина
г. Выгодное, но не обязательное совместное существование А
59. В естественном биогеоценозе взаимодействуют факторы:
а. Биотические
б. Абиотические и биотические
в. Биотические и антропогенные Б
60. Какой из компонентов экосистемы наиболее подвержен изменениям?
а. Продуценты
б. Консументы
в. Редуценты Б
61. Степные экосистемы сформировались под влиянием:
а. Наличия грунтовых вод недалеко от поверхности
б. Сухого климата и диких копытных животных
в. Хозяйственной деятельности человека Б
62. Какое количество энергии переходит на каждый последующий уровень?
а. 1 %
б. 10 %
в. 25 % Б
63. Какой фактор экосистемы смешанного леса является ограничивающим?
а. Вода
б. Пища
в. Почва Б
64. Сообщество растений называется
а. Фитоценоз
б. Зооценоз
в. Биоценоз А
65. Выпадение какого звена в цепи питания может привести к серьезным последствиям для биоценоза?
а. Редуценты
б. Консументы 2
в. Консументы 1 А
66. Выбери экосистему, в которую не включены автотрофные организмы
а. Пустыня
б. Пещера
в. Тундра Б
67. Найди правильную цепь питания сообщества тундры:
а. Растение - мышь - песец - сова
б. Мох - олень - сова
в. Карликовая береза - гриб-трутовик - олень А
68. Найди правильную цепь питания сообщества пруда:
а. Бактерии - простейшие - мелкие рачки - рыба - хищная рыба
б. Водоросли - инфузория туфелька - дафнии - водяной клоп - личинка стрекозы
в. Водоросли - перловица - ракообразные В
69. Известно, что продуктивность экосистемы зависит от лимитирующих факторов. Назовите лимитирующий фактор для лесной зоны:
а. Количество осадков
б. Количество тепла
в. Содержание в почве питательных элементов В
Библиография
1. Агесс П. Ключи к экологии, Пер.с фр., Л , 1982
2. Андерсон Дж.М. Экология и науки об окружающей среде: биосфера, экосистемы, человек, Л., Гидрометеоиздат, 1985
3. Будыко М.И. Глобальная экология, М., Мысль, 1977
4. Введение в экологию, Под ред. Ю.А. Казанского, М., ИздАт, 1992
5. Козлова Т.А., Сухова Т.С., Сивоглазов В.И. Экология, Книга для учителя, М., Школа-пресс, 1996
6. Криксунов Е.А., Пасечник В.В. Экология: Тематическое планирование, 9 класс, М., Дрофа, 1995
7. Рамад Ф. Основы прикладной экологии: Воздействие человека на биосферу, Пер.с фр., Л., 1981
8. Ревелль П., Ревелль Ч. Среда нашего обитания, В 4-х кн., М., Мир, 1995
9. Реймерс Н.Ф. Природопользование: Словарь-справочник, М., Мысль, 1990
10. Реймерс Н.Ф. Охрана природы и окружающей человека среды: Словарь-справочник, М., Просвещение, 1992
11. Стадницкий Г.В., Родионов А.И. Экология, М., Высшая школа, 1988
12. Суравегина И.Т., Мамедов Н.М. Экология: Задания, тесты, Рабочая тетрадь, М., Школа-Пресс, 1996