- Преподавателю
- Биология
- Конспект урока биологии в 10 классе Состав и строение биосферы
Конспект урока биологии в 10 классе Состав и строение биосферы
| Раздел | Биология |
| Класс | 10 класс |
| Тип | Конспекты |
| Автор | Уткина А.С. |
| Дата | 15.11.2015 |
| Формат | doc |
| Изображения | Есть |

Состав и строение биосферы.
Термин биосфера был предложен в 1875 г. австрийским геологом Зюссом. В начале XX в. Вернадским было разработано учение о биосфере. Согласно Вернадскому биосфера - это оболочка Земли населенная живыми организмами, активно ими преобразуемая. Биосфера - это экосистема глобального масштаба. Поскольку экосистема и биогеоценоз являются синонимами, то и биосферу можно рассматривать как глобальный биогеоценоз. Поэтому раздел экологии изучающий функционирование биосферы относится к биоценологии. Биоценология изучает структурно-функциональную и пространственно-временную организации экосистем различных масштабов. Задачи биоценологии состоят в изучении общих закономерностей, строения, функционирования, распределения, образования и эволюции экосистем.
Биогеоценоз включает в себя две равноценные и взаимодействующие компоненты: биогеоценоз и биотоп.
Изначально с момента возникновения жизни на Земле эти компоненты были так или иначе взаимосвязаны в составе биосферы и изменялись совместно.
Рассматривая эволюцию биосферы, биологи традиционно в центр внимания ставят живую компоненту, а геологи физико-химическую основу. Не живая компонента имеет более раннюю историю, она существовала и развивалась за долго до того как появилась жизнь. Формирование и эволюция жизни протекала при постоянном взаимодействии со средой. Количество химических элементов вовлеченных в процессы функционирования самоорганизующейся жизни постоянно увеличивалось, более 40 химических элементов вошли в сложные соединения прочно связанные с живой массой. В результате развития жизни и некоторых других физико-химических биогеохимических процессов современная среда обитания организмов принципиально отличается от исходной. Совместная эволюция живой и неживой материи породила представление о Гее - о биосфере как о сверхорганизме эволюционирующим в направлении максимальной комплиментарности живой и неживой материи.
Невозможно представить себе существования жизни вне среды обитания, вне вещества и энергии. Вещество, энергия и информация буквально «текут» сквозь организмы и этот поток обеспечивает существование жизни. По сути дела среда является составной частью жизни, т.е. жизнь - это физико-химико-биологическая система, которая не может быть отделена от единственно известной формы существования экосистемы.
Важно учитывать, что наблюдаемое сообщество и экосистемы - это не окончательно сформировавшаяся структура, а тонкие временные срезы в длительном историческом процессе их развития. Чем крупнее экосистема, тем более замкнутые круговороты вещества в ней и тем выше её способность к самоподдержанию. О продолжительности времени существования экосистем можно судить по времени оборота вещества, это максимальное физическое время, в течение которого проявляются все свойства экосистем как целого. В биосфере время оборота вещества составляет более 100 млн. лет. Цикл определяется тектоническими процессами, ведущими к горообразованию и эрозии горных пород с последующим возвращением их с речным стоком в океан. В элементарных экосистемах таких как, например «черные курильщики», заливы, лагуны время оборота основного вещества составляет от нескольких десятков до сотен лет и больше. Самая целостная их известных систем - это биосфера. Её можно рассматривать как мозаику из произвольного количества экосистем меньшего масштаба адаптируемого к конкретным геохимическим условиям. Различия между экосистемами в размерах занимаемого пространства, сложности самих систем, замкнутости биогеохимических круговоротов и продолжительности их самостоятельного существования.
Природные экосистемы - это сложно структурированное образование. Структура экосистем определяется и поддерживается её функциями и многочисленными связями с другими системами, через поток энергии и миграцию элементов. Основные функции экосистем: продуцирование, потребление и разложение органического вещества, благодаря чему осуществляется связь между потоком энергии и транспортом вещества. Функционирование взаимозависимых компонентов экосистемы в процессе преобразования энергии, вещества и информации обуславливает её целостность.
Можно различить внутреннюю и внешнюю составляющую функционирования экосистемы. Внутренняя направлена на текущие процессы в данной экосистеме и поддержание целостности. Внешняя на взаимодействие с другими смежными экосистемами и поддержании целостности структур более высокого ранга вплоть до биосферы.
В современной литературе встречается два главных методических подхода, т.е. два направления изучающих жизнь на Земле: биологический, отдающий приоритет свойствам организма, в основе этого подхода лежат представления об эволюции путем естественного отбора. В этом случае изучается жизнь сама по себе, механизмы её эволюции, включающие изменчивость, наследственность и отбор. Среда выступает в качестве фактора изменчивости и отбора. Второе направление биоценотическое, отдающее приоритет взаимодействию между живыми и не живыми энергетическими системами. При этом подходе организмы наряду с физической природой являются равноценными участниками биогеохимических превращений в экосистемах. Эволюция видов рассматривается как совместная эволюция - коэволюция, всего связанного с жизнью организмов и обеспечивающего биогеохимические циклы. На каждом уровне организации жизнь имеет свои специфические задачи и способы их решения, которые гарантируют надежность существования.
Для организма принципиально важно выжить, для популяции увеличить собственные массы и территории, для сообщества найти баланс интересов популяций разных видов, для экосистемы обеспечить наиболее эффективное использование живых и неживых ресурсов.
В организации живой и неживой природы присутствует иерархичность и детерминирование.
По Вернадскому в биосфере выделяется несколько типов веществ:
- живое (биомасса всех живых организмов);
- биогенное (созданное живыми организмами, например, нефть);
- косное (вещество, образованное без участия живых организмов);
- биокосное (созданное одновременно и живыми и неживыми организмами).
Живые организмы играют важную роль в биосфере. Они являются мощным геохимическим и энергетическим фактором. К основным функциям живого вещества относятся: газовая функция (поддержание атмосферных свойств за счет фотосинтеза и дыхания), концентрационная (способность живых существ активно поглощать и накапливать определенные элементы), окислительно-восстановительная способность.
Все живые существа распределены в пределах биосферы неравномерно.
Современная наука о биосфере - это системная дисциплина, объединяющая данные биологии, экологии, геологии, химии, климатологии, почвоведения и ряда других наук.
Состав и строение биосферы.
Живые организмы обогащают окружающую среду кислородом, регулируют количество углекислого газа, солей различных металлов, т.е. поддерживают необходимый для жизни состав атмосферы, литосферы и почвы. Во многом благодаря живым организмам биосфера обладает свойством саморегуляции. Огромная средообразующая роль живых организмов позволило ученым, создать гипотезу о том, что почва создана самими живыми организмами в процессе эволюции, а современным составом атмосферы мы также обязаны живым организмам. Энергетической основой существования жизни является Солнце, поэтому биосферу можно определить как живую оболочку Земли, свойства и структура которой формируется совместной деятельностью живых организмов и определяется постоянным притоком солнечной энергии. Общеизвестно, что жизнь сосредоточена в трех традиционно выделенных геосферах: атмосфера, гидросфера, литосфера. Они соответственно подразделяются на биосферу: это аэробиосфера (населенная аэробионтами), гидробиосфера и литобиосфера. Обращает на себя внимание тот факт, жизнь сосредоточена не только на земной поверхности, но и внутри. Биоценозы над водой и на ней, в пленке поверхностного натяжения практически отсутствует. Организмы либо внутри водные, либо сухопутно-водные, но не строго воздушно водные как на границе сред на суше. На границе сред на суше обычно формируется наиболее продуктивный слой который называется фитосферой. Этот слой формируется толщей наземной растительности с сопутствующими представителями других царств. Фитосфера вместе с фотосферой в океане составляют активную пленку, но фитосфера не испытывает той биосферной структуры которая системно образовалась на поверхности суши - террабиосфера. В террабиосферу входят гетеротрофные эоловые зоны высокогорий.
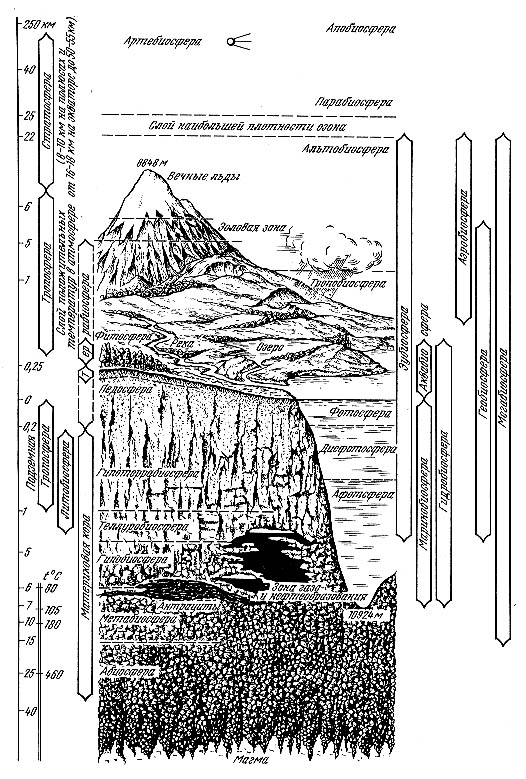
Строение биосферы
Фитосфера и эоловые оны составляют биогеоценотический покров, однако жизнь не ограничивается лишь поверхностью суши. Она проявляется, хотя и не столь явно, как на поверхности, в глубинах литосферы и особенно в подземных водах - телуробиосфера. Но под педосферой расположен слой интенсивноговыветривания котоый называется гипотеррабиосфера. Раздел между гипотеррабиосферой и телуробиосферой лежит на кислородной границе (ниже 1 км), ниже которой подземные воды не содержат свободного кислорода. Жизнь может существовать, хотя бы теоретически, и глубже. В гидротермах дна океана («черных курильщиках») на глубинах в 3 км и давлении около 300 атмосфер были обнаружены организмы, живущие при 250° С. Вода тут не кипит из-за высокого давления (пределы жизни ограничены точками превращения воды в пар и сворачивания белков). Перегретая жидкая вода обнаружена в литосфере до глубины 10,5 км, так что теоретически там могут существовать и организмы. Глубже 25 км, по оценкам, должна существовать критическая температура в 460° С, при которой при любом давлении вода превращается в пар, и жизнь невозможна. Случайно попавшие в глубь земных пластов организмы, уже не способные к активному метаболизму из-за высоких температур и огромного давления горных пород, но живые - в виде спор и цист, составят гипобиосферу - аналог парабиосферы в атмосфере (см. ниже). Глубже расположен слой эволюционно биогенной литосферы без признаков современной активной жизни - метабиосфера.
Ниже гранитного слоя нет ни современного, ни геологически прошлого воздействия жизни или оно нацело стерто глубинными процессами. Тут расположена абиосфера планеты как космического тела.
Продвигаясь вверх от поверхности Земли, как это было только что проделано при движении в ее глубины, обнаруживаем весьма разреженную жизнь аэробиосферы, где в нижнем слое тропобиосферы она представлена не только микроорганизмами и вирусами, главным образом живущими в капельках атмосферной влаги, но довольно многочисленными представителями животного и растительного миров, поднимающимися с поверхности земли и растительного покрова. В основном большую часть организмов надо отнести к экотону между террабиосферой и гидробиосферой с одной стороны и тропобиосферой с другой. Обычно граница между экотоном и собственно тропосферой проходит на высотеот 50 до 300 м. Над тропосферой вне слоя положительных температур лежит относительно тонкий слой альтобиосферы (лат. altus - высокий). Он приблизительно соответствует эоловой зоне на суше. Здесь жизнь возможна лишь благодаря прямой солнечной инсоляции, тогда как температура среды не поднимается выше 0° С.
Над альтобиосферой расположен слой случайного заноса организмов и их спор - парабиосфера. Над паробиосферой лежит слой апобиосфера, т.е. слой без признаков жизни.
В атмосфере пределы жизни - 8-17 км.
В гидросфере - до 11800 км.
В литосфере - 5-6 км.
На суше толщина активного слоя жизни порядка 12 км, а в пределах океана порядка 17 км.
Горизонтальная структура биосферы

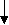
 БИОСФЕРА
БИОСФЕРА
аэробиосфера геобиосфера
гидробиосфера 

 террабиосфера литобиосфера
террабиосфера литобиосфера
т ропобиосфера
ропобиосфера
марино- фотосфера аква- педосфера
а льтбиосфера биосфера биосфера
льтбиосфера биосфера биосфера
гипотерра-
афотосфера биосфера
 царство
царство
область теллуро-
биозона биосфера
биом
биолокус
биокомплекс
биогеоценоз
Высший уровень деления террабиосферы - биогеографическое царство. Для следующего более низкого системного уровня пока нет устойчивого названия. Обычно это материковый блок, океан или их крупные части. Ведущий системообразующий фактор тут, главным образом, эволюционно-исторические условия и события формирования биоты, ее взаимодействия со средой. Это взаимодействие прежде всего связано с энергетикой процессов, и потому области распадаются на природные пояса (в биосферной системе терминологии - биозоны), в рамках которых история и форма биотического обмена на суше формируют биомы. Внутри биомов, или биозон, направление сукцессионных процессов, определяемых литогенной основой и местными особенностями климата и почвообразования, создает индивидуальный ландшафт или биолокус (от лат. loca - ландшафт). В пределах биолокусов благодаря взаимосвязям вещественно-энергетических и информационно-экологических компонентов формируются биогеоценозы, а точнее биоценотические комплексы. Самые малые круги биоценотического обмена на основе относительной гомогенности территории формирующей экосистемы или биогеоценозы.
Таким образом, предполагается, что в биосфере имеется по крайней мере 8-9 уровней относительно самостоятельных круговоротов веществ, в пределах взаимосвязи 7 основных вещественных, энергетических и экологических компонентов и 8 информационного.
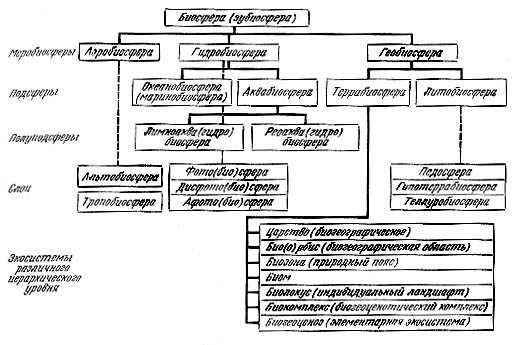
Иерархия экосистем биосферы
Субстратом в аэробиосфере и гидробиосфере служит вода (газы, видимо, принципиально не могут служить субстратом для жизни). Включение энергетики как экологического компонента достаточно обосновано тем, что все процессы в природе по своей основе всегда энергетические. Экологически переработка информации - также неотрывное свойство живого. Глобальные, региональные и местные круговороты веществ незамкнуты и в рамках иерархии экосистем частично «пересекаются». Это вещественно-энергетическое и отчасти информационное «сцепление» обеспечивает целостность экологических надсистем вплоть до биосферы. Природные системы океана и суши относительно биотически не зависимы. Однако их геосистемы как биотопы очень тесно взимосвязаны. Однако в основном все-таки взаимодействуют биоты аквабиосферы и маринобиосферы, т.е. гидробиосфера и геобиосфера в значительной мере автономны как биотические образования. Геохимический сток с суши в океан практически однонаправлен. Эволюционно гидробиосфера и геобиосфера развиваются практически независимо друг от друга.
Для биосферы в целом на ее входе имеется энергия, земное и космическое вещество, на выходе - осадочные биогенные породы и уходящие в космос газы.
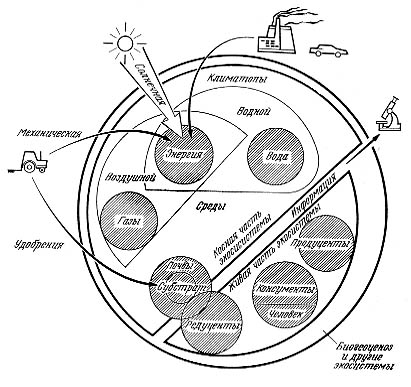
Экологические компоненты
Общие закономерности организации биосферы Земли.
Современная экология исходит из аксиомы иерархической организации или принципа интегративных уровней, с учетом аксиомы эмерджентности. При этом справедлив закон преломления космических воздействий, т.е. космические факторы оказывают влияние на биосферу и её подразделения при этом подвергаются изменению со стороны экосфер Земли и по этому по силе и времени проявления могут быть ослаблены, сдвинуты или вовсе утратить свой эффект. Системы биосферы могут блокировать космические воздействия либо целиком, либо частично. Биосферу, как и любую другую систему, формируют и не только и не столько внешние факторы, а внутренние закономерности. Одним из важнейших свойств биосферы является закон биогенной миграции атомов открытый Вернадским «Миграция химических элементов на земной поверхности и в биосфере в целом осуществляется при непосредственном участии живого вещества (биогенная миграция) или протекает в среде, геохимические особенности которой обусловлены живым веществом, как тем которое в настоящее время населяющих биосферу, так и тем, которые действуют на Земле в течение истории».
Согласно этому закону понимание общих химических процессов на поверхности планеты не возможно, без учета биотических и биогенных факторов, в том числе и эволюционных. Но поскольку человечество сильно влияет на биоту, они тем самым изменяют условия биогенной миграции атомов, что может привести к глубоким химическим сдвигам в перспективе, поэтому процесс может стать саморазвивающимся и практически не управляемым (например, опустынивание). Особенности биогенной миграции атомов связаны со степенью замкнутости глобальных биогеохимических круговоротов, поэтому существует правило не замкнутости биотических круговоротов (если круговорот был бы замкнут, то не могла бы происходить эволюция).
Количество живого вещества в рамках больших геологических отрезков времени не подвержена заметным изменениям, эта закономерность была сформулирована в виде закона константности количества живого вещества (Вернадский). «Количество живого вещества для данного геологического периода остается постоянным».
Как аккумулятор солнечной энергии живое вещество одновременно реагирует как на внешние космические изменения, так и на внутренние. Увеличение или снижение количества живого вещества в одном месте биосферы должно приводить к синхронному процессу с обратным знаком в другом регионе. Однако следует учитывать скорость процесса, если человек прямо нарушает природу изменения будут выше. Кроме того не всегда происходит адекватная замена.
С законом физико-химического единства живого вещества на прямую связан закон сохранения структуры биосферы. Для сохранения структуры биосферы живое вещество стремится к достижению состояния зрелости или экологического равновесия. Единство живого вещества и гомологичность строения подсистем биосферы приводит к тому, что реализуется принцип гетерогенезиса живого вещества. Гетерогенезис подчиняется принципам экологической комплементарности и принципу экологического порядка, следствием этого является стабильность всей системы биосферы, заполнение пространства внутри природных систем упорядочено таким образом, что позволяет реализоваться гомеостатическим свойствам системы с минимальными противоречиями внутри её (закон упорядоченности заполнения пространства и пространственно-временной определенности). С этим следствием на прямую связан принцип системной дополнительности, т.е. подсистема одной природной системы в своем развитии обеспечивают предпосылку для успешного развития и саморегуляции других подсистем входящих в эту систему (если она не подвергается мощной внешней деформации).
Правило автоматического поддержания глобальной среды обитания: «Живое вещество в ходе саморегуляции и взаимодействия с абиотическими факторами автодинамически поддерживает среду жизни пригодную для её развития».
Закономерности развития эволюции биосферы.
Эволюция живого началась с возникновения форм пред-жизни, а затем и проорганизмов, с этого геологического «момента» начал действовать принцип Реди: «живое происходит только от живого, между живым и не живым веществом существует граница, хотя и имеется постоянное взаимодействие». Этот принцип служит основой формирования экосистем. Понятно, что геологические процессы и явления с течением времени изменялись, поэтому геоэкологическую историю трудно смотреть с современной точки зрения. Для биосферы любого этапа развития характерны круговороты веществ. С течением времени увеличивается доля биологического компонента в замыкании биогеохимического круговорота.
Человеческая деятельность ведет к гомогенизации систем биосферы, при этом снижается степень замкнутости биогеохимических циклов, а следствием этого является загрязнение.
Саморазвитие биоты шло двумя путями: по внутренним законам и согласно внешним воздействиям с адаптацией к ним. Это закон эколого-системной направленности эволюции, чем больше организмы воздействуют на биосферу, тем интенсивнее идет их эволюция. Этот принцип максимального эффекта внешней работы или закон исторического развития биологических систем был сформулирован Бауером в 1935 г: «Развитие биологических систем есть результат увеличения их внешней работы, т.е воздействие систем на их окружающую среду».
Физико-математическая интеграцию этого принципа дает теория порога возрастающей энтропии в биосфере. Эту теорему сформулировал Тринчер в 1964 г. Теорема порога возрастающей энтропии как бы противоположна теореме о сохранении упорядоченности в открытой системе. Связано это с довольно вольным значением понятия энтропия. Если определить его как меру вероятности пребывания системы в данном состоянии (согласно принципу Больцмана о переходе системы из менее вероятного состояния, в более вероятное, т.е. равновесное) и признать жизнь как неравновесное (квазиравновесное) состояние систем, то падение энтропии или её возрастание до порогового уровня означает практически одно и то же, т.е. энтропийные рамки жизни. Важно лишь то, что минимум энтропии возникает при не равновесном распределении вещества в системе. Человеческая деятельность нарушает эту неравномерность, делает живое вещество гомогенным, видоизменяет энтропийные и энгтропийные процессы.
Антропогенное воздействие на окружающую среду оказывает деструктивное влияние, оно заменяет биогенную эволюцию разрушая, естественные системы природы. Эволюция вынуждена идти экстенсивно под воздействием внешних факторов с темпом диктуемым трансформированием природы человеком, а не ходом естественных явлений.
Закон исторического развития биосистем уже не работает или работает не в полной мере, т.к роль биотических воздействий на среду относительно снизилась. Доминирует преобразующая деятельность человека. В следствии за прямым уничтожением видов возможна самодеструкция всего живого. Фактически этот процесс и идет в виде массового размножения отдельных организмов разрушающих сложившиеся экосистемы.
Для биосферы наиболее опасным является ускорение темпов изменений. Сама эволюция биосферы не была равномерной, степень совершенства биогеохимического круговорота постоянно увеличивалась, но этот процесс не шел гладко. Широко известны эволюционные катастрофы, отмеченные в истории планеты на рубежах 650, 450, 230, 65 млн. лет назад. Наиболее древний эволюционный кризис привел к «внезапному» исчезновение многих видов одноклеточных водорослей.
На рубеже 450 млн. лет вымерло большинство плацентарных обитателей океана, на следующем этапе ускорения эволюционных процессов на рубеже 230 млн. лет исчезли большинство видов гигантских амфибий. 65 млн. лет назад довольно быстро вымерли гигантские рептилии и многие другие группы организмов. Этот последний этап эволюционных смен привлекает наибольшее внимание. Его связывают с падением на Землю огромного метеорита-астероида, предположительно образовавшего самый гигантский кратер на территории современной Мексики. Ход вымирания не достаточно ясен. Предполагается похолодание, которое привело к нарушению инкубации яиц рептилий и т.п. аномалиям. Механизмы вымирания могли быть и другими. Весьма незначительные изменения абиотической среды ведут к едва заметным, на взгляд человека, энергетическим перестройкам. Вид никогда не исчезает один. Всегда сложно изменяются пищевые и информационные сети, происходит каскадное по иерархии систем глобальная перестройка. Одни виды безвозвратно исчезают, другие в силу энергетических правил, в том числе и необходимости каскадного преобразования солнечной энергии их замещают. Это убыстрение отражено в виде правила катастрофического толчка: природная или природно-антропогенная катастрофа всегда приводит к существенным природным перестройкам, которые относительно прогрессивны для биосферы, т.к. адаптируют её системы к новым условиям среды.
Поскольку существует ускорение и замедление эволюции, действует принцип прерывности и непрерывности развития биосферы: процесс медленного эволюционного изменения организмов закономерно прерывается фазами бурного развития и вымирания практически безпереходных форм. Равномерно-линейная дарвинская эволюция математически не реальна. Эволюция явно шла по касс каскадному принципу. Действовал лингвистический принцип хорошо освещенный в работах Школя для генетического уровня биосистем: шел отбор не бук и слов, а предложений. В этом случае естественный отбор идет среди природных систем, требуя гораздо меньше времени на эволюционирование.
Математические расчеты общего разнообразия генетического кода позволили немецким биохимикам рассчитать его возраст 3,8 ± 0,6 млрд. лет. Это число вполне укладывается в современные рамки палеонтологических находок. Органическое вещество современного типа появилось 3,8 млрд. лет назад (Гренландия), а организмы 3,5 млрд. лет назад (Австралийские находки).
Биогеоиерархический принцип в эволюции мог вызвать ускорение в её ходе. Не исключено что оно было настолько велико, что промежуточных форм практически не существовало. Если ведущую роль в отборе играли верхние уровни иерархии и они же направляли эволюцию, то антропогенные изменения биосферы, идущие с большой скоростью в любой момент могут дать толчок для ускорения эволюционных перестроек. Это будет означать капитальную перестройку экологических условий на планете.
Необходимо иметь какие-то количественные данные хотя бы самые общие для выяснения, что опасно, а что еще нет в ходе ускоренной эволюции среды. Такими критическими рамками видимо могут быть «точки Пастера» и правило 1 и 10 %. Основой «точки Пастера» служит момент, когда уровень содержания кислорода в атмосфере Земли достигает примерно 1 % от современного. С этого времени стала возможной аэробная жизнь. Геохронологически - это архей. Предполагается, что накопление кислорода шло взрывообразно и заняло в эволюции не более 20 тыс. лет. Вторая «точка Пастера» - достижения содержания кислорода в атмосфере около 10 % от современного. Это привело к возникновению предпосылок формирования озоносферы, жизнь стала возможной сначала на мелководье, а затем на суше. Произошло это в архее и видимо столь же геологически стремительно.
«Точки Пастера» как и закон пирамиды энергии Лендемана дали повод для формулировки правил 1 и 10 %. Число 1 % возникает из соотношения возможности потребления энергии и «мощностей» необходимых для стабилизации среды. Для биосферы доля возможного потребления общей первичной продукции не превышает 1 %, т.е. около 1 % чистой первичной продукции в энергетическом выражении потребляют позвоночные животные как консументы высших порядков. Около 10 % беспозвоночные, как консументы низших порядков и оставшаяся часть потребляется бактериями и грибами сапрофагами. Как только человечество стало использовать большее количество продукции биосферы, так и перестал удовлетворяться принцип Ле-Шателье-Брауна (на настоящий момент человечество потребляет не менее 10 % от общей энергии биосферы, т.е. растительность не давала необходимый прирост биомассы).
Эмпирический порог потребления, приводящий к заметным изменениям в системе природы, принят на уровне 5-10 % от суммы вещества.
Ориентировочно можно разделить переходы для природных систем с организменным типом управления с одной стороны и популяцией с другой. Для первых порог выхода из стационарного состояния до 1 % от потока энергии, а порог разрушения около процента от этой нормы. Для популяционных систем нарушение в среднем 10 % объема изъятия приводит к выходу этих систем из стационарного состояния. Видимо для глобальных энергетических систем выход из стационарного состояния происходит в рамках 0,1-0,2 % от потока энергии, т.е. значительно раньше, чем начинается сбой в действии принципа Ле-Шателье-Брауна.
Эволюционные переходы в биосфере занимают сравнительно очень не большое время. Так предельная совокупность видов могла быть образована в эволюции всего за 70 млн. лет, при этом дальнейшее нарастание числа видов практически исключено, что следует из ограниченности числа экологических ниш, а соответственно и принципу конкурентного исключения. В течение последующих 0,5 млрд. лет число видов оставалось примерно одинаковым а, следовательно, существует правило константности числа видов в ходе стационарной эволюции биосферы: в среднем число нарождающихся видов в среднем равно числу вымерших. И общее видовое разнообразие константно. видимо это правило справедливо уже для сформировавшейся биосферы и возможно ограничено временем геологических периодов, логически оно вытекает из необходимости материала для экологического дублирования без которого структура биосферы теряла бы своё свойство надежности. Отсюда следует правило обязательности заполнения экологических ниш.
Практическое значение этих закономерностей было сформулировано следующим образом: другого устойчивого состояния биосферы не существует и при сохранении или росте современного антропогенного возмущения, устойчивость окружающей среды будет разрушена. Это подтверждается и правилом усиления интеграции биологических систем Шмальгаузена: биологические системы в процессе эволюции становятся все более интегрированными со все более развитыми регуляторными механизмами обеспечивающими такую интеграцию. Конкурентное взаимодействие включает в себя резные типы воздействий, формирует не только экосистемы согласно принципу комплементарности (дополнение), экологической конгруэнтности (соответствие) и закону формирования экосистем на базе соответствия биотопа и биоценоза, но и компенсируя процессы на уровне экосистем одного уровня иерархии. Кроме самых верхних уровней всегда наблюдается некий набор конкурирующих экосистем.
Дезинтеграция (разъединение) сообществ и нарушение их иерархии под воздействием антропогенных причин ведет к деструкции природных систем. Разрушение более трех уровней иерархии экосистем абсолютно не обратимо и катастрофично.
Множественность конкурентно-взаимодействующих экосистем обязательна для поддержания надежности биосферы, именно так шла её эволюция. Антропогенное воздействие нарушает этот ход. Интеграция захватывает не только биологические образования, но и многие другие с интеграцией систем увеличивается ценность информации.
Понятие системы
Система - это любое множество элементов, которые определенным образом взаимосвязаны друг с другом, т.е. система - это совокупность взаимодействующих между собой относительно элементарных структур или процессов объеденных в целое выполнением общей функции, не сводимой к функциям её компонентов.
По сложности системы разделяют на простые динамические системы с не большим количеством элементов; сложные поддающиеся описаниям; очень сложные системы, которые нельзя описать поэлементно и предсказать их эволюционирование.
По степени определенности функционирования:
- детерминированные (это такие системы, в которых элементы однозначно взаимодействуют строго определенным образом; поведение такой системы можно точно предсказать в любой момент).
- вероятностные (это системы, в которых элементы находятся под влиянием большого количества воздействий, поэтому взаимодействие всех элементов не может быть точно описано и в какой-то степени становится неопределенным; поведение такой системы трудно предсказать с достоверной точностью).
Вероятность не противопоставляется детерминированности в смысле причинности, т.е. все явления природы имеют свои причины, т.е. детерминированы.
Информация - это способ организации вещества и энергии в систему.
Энергия - это способность совершать работу.
В зависимости от характера обмена энергии и массы с окружающей средой различают три группы систем:
1. изолированные;
2. замкнутые;
3. открытые.
Процессы, которые протекают в системах, могут быть равновесными и неравновесными:
- равновесные протекают в системе таким образом, что вызванные ими изменения в состоянии системы могут пойти в обратной последовательности без дополнительных изменений в окружающей среде.
- неравновесные процессы в основном протекают в одном направлении, а протекание таких процессов в обратном направлении всегда сопровождается изменениями в окружающей среде.
Экосистема и вещество в экосистеме.
Вещество понимается как составная часть материи, часто встречаются принципиальные неточности, приравниваются понятия вещество и материя. Материя - это общее понятие, а вещество - это одна из форм существования материи. Другая форма существования материи называется физический вакуум. вещество выступает в двух ипостасях: в виде реальных полей и в виде соответствующих этим полям также реальных дискретных частиц, т.е. вещество дуалистично. Природа физического вакуума и его строение изучено намного хуже, чем вещество. Вакуум - это нулевые, флуктуирующие поля, с которыми связаны виртуальные частицы. Вакуум - это не пустота - это материальный фон, обладающий своеобразными свойствами. В квантовой теории, вакуум - это пространство, в котором отсутствуют реальные частицы, и выполняется условие минимума плотности энергии в данном объеме.
Вещество в экосистеме выступает в качестве массы, точнее количества и состава, т.е. качества. В качестве массы оно выступает, как физическая субстанция, а в качестве состава она выступает как химическая субстанция с соответствующими свойствами. Поведение живого вещества во многом наследуется поведением не живого вещества. После большого взрыва произошедшего около 15-20 млрд. лет назад, первоначальная масса непрерывно расширяется, т.е. увеличивается в объеме и распространяется в космическом пространстве.
В еще большей мере свойствами к неудержимой экспансии обладает живая масса, что обнаруживается в индивидуальном росте и размножении организмов. Однако живая масса в биосфере потенциально ограничена пространством Земли и её ресурсами. Поэтому большее количество и разнообразие живой массы, в принципе не беспредельно. Увеличение разнообразия организмов ведет к росту дискретности биомассы, т.к. биомасса распределена между особями и к возможной потере экосистемы и биосферы своей целостности. Это вводит дополнительные ограничения на количество живого вещества в биосфере. Достигнув своего физического предела, биомасса может только самообновляться, и перераспределятся между организмами. Последние около 100 млн. лет суммарная биомасса организмов постоянна, что указывает на достижение ею предела насыщения для данных условий. Принципиальная особенность поведения вещества в экосистемах связана с его обращаемостью, т.е. многократным использованием организмами в условиях лимитирования его общего количества физическими размерами планеты.
Элементы циркулируют, переходя из одной формы в другую и, в конце концов, возвращаются в исходное состояние, но с неминуемыми потерями. Жизнь на Земле тесно связана с доступностью важнейших химических элементов. В первую очередь: углерод, азот, фосфор, кальций, железо и около десятка микроэлементов. Всего жизнь использует не многим более 40 элементов. Соотношение атомов трех первых элементов в Мировом океане оцененное по океаническому планктону составляет 100:15:1, а в биосфере 110:16:1. Все используемые живыми организмами элементы включены в биогеохимические циклы, в которых они проходят сложную цепь превращений. Циклами затрагивается только часть содержащегося в земной коре и недрах Земли химического элемента. Отсюда деление фонда элементов на резервный (большой) и обменный (малый), между которыми существует слабый по интенсивности обмен, обеспечивающий соответствующее увеличение обменного фонда по мере роста общей биомассы организмов.
Циклами связаны, хоть и не жестко, многие организмы. Это придает определенную функциональную целостность экосистем. Циклы разных химических элементов не соподчинены, т.е. они не зависимы и соприкасаются с другими только через общие для них организмы. В результате в экосистемах образуются сложные системы циклов.
Эффективность возврата элементов может служить показателем вещественной целостности экосистем. Жизнь несет функцию эффективного превращения элементов в биогеохимических циклах и поэтому её эволюционный уровень может оцениваться с позиции достижения совершенства биогеохимических циклов. Большинство изученных биогеохимических циклов оказываются далекими от совершенства даже в масштабе всей биосферы. Они не являются до конца замкнутыми, т.к. часть вещества выходит из круговоротов в виде ископаемых и глубоководных осадков. И лишь по прошествию длительного времени в той или иной мере компенсируется вулканическими или глубинными выбросами. Степень замкнутости биогеохимических циклов зависит от меры участия в них живых организмов, а также вовлеченности организмов в трансформацию вещества и их количественное и видовое разнообразие.
В масштабе биосферы наиболее совершенным является цикл азота, который включает в себя процессы трансформации вещества, охватывающие атмосферу, литосферу и гидросферу.
Основной резервуар воды сосредоточен в литосфере в связанном состоянии (250 тыс. геограммов). В океане сосредоточено 13 800 геограммов, в ледниках 167, грунтовые воды 2,5, внутренние воды 0,25, в атмосфере 0,13. Особенно важны 2-а аспекта круговорота воды:
1. Море теряет из-за испарения воды больше, чем получает её с осадками. На суше положение обратное, т.е. наземные экосистемы поддерживаются за счет испарения с поверхности океана.
2. Вес воды в континентальных водоемах соответствует годовому стоку, т.е. время оборота составляет около 1 года (вес воды в континентальных водоемах 0,25, а годовой сток 0,2 геограмма). Разность между количеством осадков за год (1 геограмм) и стоком (0,2 геограмма), совместно с испарением (0,6 геограммов) составляет 0,2 геограмма.
О - С - И = 1 - 0,2 - 0,6 = 0,2
Эта цифра соответствует величине годового поступления воды в подпочвенные водоносные горизонты. Увеличение стока в результате деятельности человека приводит к уменьшению очень важного для круговорота фона грунтовых вод и может быть нивелировано (выровнено); только правильное хранение воды в подземных резервуарах, а не в озерах и водохранилищах, откуда она быстро испаряется.
Основной резерв углекислого газа в океане находится в виде карбонатов (130 тыс. млрд. тонн), а также в литосфере в виде горючих ископаемых 40 тыс. млрд. тонн, и в атмосфере 2 300 млрд. тонн. Только малая часть углекислого газа (1,3 %) вовлекаются в биогеохимический круговорот. Фотосинтезирующие организмы и карбонатные системы моря весьма эффективно удаляют углекислоту из атмосферы, однако потребление человеком горючих ископаемых вместе с уменьшением поглотительной способности фотосинтетиков начинает влиять на атмосферный фон круговорота.
Можно ожидать, что в ближайшее время устанавливается новое, но ненадежное равновесие между постоянно увеличивающейся концентрацией углекислоты (15 % за последние 100 лет) и пылевых загрязняющих частиц, которые являются физическими барьерами для космического и теплового излучения. Биосферный круговорот других веществ и элементов изучен в основном на качественном уровне (пример, цикл серы, который включает геохимические, биогеохимические, химические и биологические процессы трансформации серосодержащих соединений). Химическая реакция окисления этих веществ сопровождается выделением энергии; некоторые экосистемы, в частности экосистемы сероводородных зон используют эту энергию для хемосинтеза или физиологических процессов. Одновременно многие бактерии восстанавливают окисленные сульфиды обеспечивая биотический баланс. Чисто биогенное происхождение имеет диметил сульфид, придающий морю его характерный запах. Он продуцируется одновременно водорослями родов, эмилиани, и фиацистис, и в большом количестве переходят из океана в атмосферу. Его количество в тропосфере определяет её облачность и т.д. эпипелагические экосистемы могут регулировать глобальный климат. Однако очень многое в биосферном цикле серы и других элементов остается не изученным, особенно количественный аспект их глобальной миграции.
Энергия в экосистеме.
Энергия - это неотъемлемое свойство материи, мера движения, степень интенсивности и способности к внешним преобразованиям. В классической физике энергия - это функция состояния системы зависящая от значений параметров давления, объема, температуры и т. д., которые определяют состояние. В закрытых и равновесных системах превращение энергии подчиняется двум законам термодинамики. Согласно второму закону рассеивание происходит от концентрированной к рассеянной форме. Это приводит к термодинамическому равновесию.
Первый закон термодинамики - это закон сохранения энергии. Энергия может переходить из одной формы в другую, но она не исчезает и создается заново.
Второй закон термодинамики устанавливает критерии, отражающие одностороннюю направленность необратимых процессов не зависимо от их конкретной природы. Согласно этому закону состояние системы может быть описано особой функцией состояния - энтропией. В изолированной системе энтропия остается не изменой в равновесных и возрастает в неравновесных процессах. Таким образом, протекающий в изолированной системе самопроизвольный неравновесный процесс всегда вызывает увеличение энтропии до ее максимальных значений при окончании процесса и установление термодинамического равновесия.
Энтропия, как функция состояния термодинамической системы отражает следующие процессы связанные с превращением энергии могут происходить самопроизвольно только при условии, что энергия переходит из концентрированной формы в рассеянную, т. е. деградирует. Рано или поздно такие процессы приводят систему к ее самому простому состоянию - термодинамическому равновесию. Это состояние характеризуется минимальной упорядоченностью. Все виды энергии деградируют переходя в тепловую, которая в среднем равномерно распределена между всеми элементами системы.
Мера, количество связанной энергии, которая становиться не доступной для использования называется энтропией. Этот же термин используется как мера изменения упорядоченности, которое происходит при деградации энергии.
Непосредственное применение второго закона термодинамики к открытым системам, в которых происходит неравновесные процессы, встречается ряд трудностей. Критерием направления самопроизвольных изменений в изолированной системе служит увеличение энтропии, а конечным состоянием равновесие. В то же время открытые системы первую очередь биологически прекращают свое функционирование в состоянии равновесия, т. е. поддержание неравновесных состояний в открытых системах возможно лишь за счет создания в них соответствующих потоков вещества и энергии. Изменение энтропии открытой системы может происходить за счет процессов обмена системы с внешней средой, либо за счет возникновения энтропии в самой системе вследствие внутренней необратимости изменений, т.е. постулируются, что общее изменение энтропии открытой системы dS складывается из двух независимых частей. Из процессов обмена системы с внешней средой плюс возникновение энтропии в самой системе вследствии внутренней необратимости изменений
dS = dèS + diS.
Если внутри системы протекают обратимые изменения, то они не сопровождаются возникновением энтропии и diS = 0.
В случае необратимых изменений diS >0.
Очевидно, что в изолированных системах где deS = 0 dS = deS + diS сводиться к dS = diS >0.
Если в каком - либо участке открытой системы одновременно протекают различные необратимые процессы, то величина diS >0 описывает приращение энтропии, которое является следствием взаимодействия необратимых процессов друг с другом. Разделение величины изменения энтропии открытой системы на две составляющие позволяет изучать термодинамические свойства открытых и изолированных систем.
Если продифференцировать уравнение, то получим
dS/dt = deS/dt + diS/dt.
Это уравнение означает, что скорость изменения энтропии системы равна скорости обмена энтропией с окружающей средой плюс скорость возникновения энтропии внутри системы.
При diS/dt>0 общая энтропия системы может как возрастать, так и убывать.
Положительная величина deS/dt связаны с увеличением энтропии системы в результате обмена веществом и энергией с внешней средой.
Отрицательная величина deS /dt < 0 соответствует тому, что отток положительной энтропии от системы во внешнюю среду превышает приток положительной энтропии из вне, так что в результате общая величина баланса обмена энтропией между системой и средой является отрицательной. При одном и том же условии при diS /dt >0 (скорость возникновения энтропии >0) возможны три следующих случая:
-
Общая энтропия системы >0 если скорость обмена энтропии средой и системы > 0 или она меньше 0, но модуль меньше чем dS/dt
dS/dt > 0 если
dS/dt > 0 или
deS/dt < 0, но
| deS/dt| < 0 diS dt
-
dS/dt < 0 если
deS/dt < 0 и
|deS/dt| > diS/dt
-
dS/dt = 0 если
deS/dt < 0 и
|deS/dt| = diS/dt
Последний случай соответствует установлению в системе стационарного состояния, при котором продуцирование энтропии в системе diS компенсируется оттоком положительной энтропии во внешнюю среду, так что общее изменение энтропии = 0
dS = deS + diS = 0
dS/ dt = deS/dt + diS/dt = 0
Анализ общих свойств биологических систем на основе дифференцированного уравнения помогает объяснить внешнее противоречие между поведением организмов и вторым законом классической термодинамики. Действительно рост и развитие организмов сопровождается усложнением организации, и сточки зрения термодинамики выглядит как самопроизвольное уменьшение энтропии живых систем, что противоречит второму закону термодинамики. Однако это противоречие лишь кажущееся, так как направление самопроизвольных процессов определяется увеличением энтропии лишь для изолированных систем, а не для не открытых, которыми являются биологические системы.
В реальных условиях развитие организмов сопровождающееся уменьшением общей величины их энтропии происходящих при условии второго случая
deS/dt < 0
|deS/dt| > diS/dt
За счет того, что в других участках внешней среду идут сопряженные процессы с образованием положительной энтропии. Общий энергообмен живых организмов можно упрощенно представить, как образование в процессе фотосинтеза сложных молекул углеводов из углекислого газа и воды с последующей деградацией продуктов фотосинтеза в процессах дыхания. Именно этот энергообмен обеспечивает существование и развитие как отдельных организмов, которые являются звеньями в потоке энергии, так и жизни на Земле в целом. С этой точки зрения уменьшение энтропии живых систем в процессах их жизнедеятельности обусловлено в конечном итоге поглощением квантов света фотосинтезирующими организмами, что однако с избытком компенсируется образованием положительной энтропии в ядерных реакциях на солнце. Этот принцип относиться и к отдельным организмам для которых поступление из вне питательных веществ несущих приток отрицательной энтропии, всегда сопряжено с продуцированием положительной энтропии при их образование в других участках внешней среды, так что суммарное изменение энтропии системе «организмы плюс внешняя среда» всегда положительно. В целом уменьшение энтропии живых систем в процессе их роста происходит за счет свободной энергии, которая освобождается при распаде поглощаемых из вне питательных веществ или за счет энергии солнца. Способность создавать и поддерживать высокую степень внутренней упорядоченности, т. е. состояния с низкой энтропией - это важнейшая термодинамическая характеристика организмов, экосистем и биомассы в целом. Низкая энтропия достигается постоянным и эффективной рассеянной и легко используемой энергии и превращение ее в энергию, используемую с трудом.
Упорядоченность экосистемы, т. е. сложная структура, достигается за счет дыхания сообщества, в результате чего энтропия диссипируется (рассеивается). Такие системы существующие и поддерживающие свою организацию, за счет использования и рассеивания энергии из внешнего источника называется диссипативной иными словами для поддержания сложности биологической системы должны использовать энергию, которую эта экосистема использовать уже не сможет. Совокупность существующих богатых энергией состояний вещества составляют энергетические ресурсы земли. Кинетическая энергия заключена в скорелированном движении различных частей вещества (солнечная энергия, энергия приливов и отливов). Потенциальная энергия содержится в скореллированных связях различных частей вещества (гравитация, ядерная энергетика, а так же скрытая теплота).
При протекании процессов распада все виды потенциальной энергии сначала переходят в кинетическую энергию упорядоченного движения, которая за этим подвергается диссипации и рассеивается в виде тепловой энергии хаотического движения молекул. Таким образом, экосистемы и организмы представляют собой открытые, не равновесные термодинамические системы, постоянно обменивающиеся с окружающей средой энергией и веществом уменьшая при этом энтропию внутри себя, но увеличивая энтропию во вне в согласии с законами термодинамики.
Связь информации с термодинамикой. Энтропия является функцией состояния системы. Увеличиваясь при любом изменении в системе она является мерой энергии, которую нельзя получить в первоначальной форме. Из второго закона термодинамики следует, что энтропия всегда возрастает в любом спонтанно протекающем необратимом процессе, которые происходят в изолированной системе. Накопление информации включает в себя энергию обмена сопровождающийся соответственным возрастанием энтропии. Информация связанная с некоторой структурой может в дальнейшем направлять энергию туда где ее использование наиболее эффективно. Сама информация тоже становиться эффективной по мере того как переходя на следующий уровень она может контролировать канал связи и организовывать вход информации, т. е. информация иерархична. Перенос энергии пищи от ее источника через ряд организмов называется пищевой цепью. В соответствии со вторым законом термодинамики поток энергии с каждой ступенью трофической пирамиды уменьшается. Принципы организации пищевых цепей связаны с действием законов термодинамики. Пищевые цепи можно разделить на два основных типа: пастбищный и детритный. Пищевые цепи не изолированы друг от друга, а тесно переплетаются образуя пищевые сети. Величины всех частей энергии чистой продукции, которые текут по двум путям в различных экосистемах разного типа и часто варьируют по сезонам или пот годам в одной и той же экосистеме. Во всех экосистемах пастбищные и детритные цепи взаимосвязаны так, что ответ на энергетическое воздействие в системе может быстро происходить переключение потоков. Не вся пища съедается пасущимися животными и усваивается, часть ее уходит в детритную цепь. Степень влияния травоядных животных на сообщества зависит не только от количества, ассимилированной ими энергии и пищи, но и от скорости изъятия животными растений . Морской зоопланктон обычно выедает большие фитопланктоны чем может ассимилировать, излишек выводиться в дитритную пищевую сеть. В наземных экосистемах дитритные цепи могут начинаться с диспергированного органического вещества или с растворенного органического вещества. Пастбищные цепи включают кроме традиционного прямого выедания зерновые пищевые цепи, которые начинаются с семян и некоторые пищевые цепи, которые начинаются с нектара цветковых растений.
В водных экосистемах поток энергии идет по четырем путям: анаэробный путь, прямое выедание растений, потребление растворенного органического вещества и потребление диспергированного органического вещества.
Источник энергии.
В большинстве экосистем суши энергия местного происхождения. Органическое вещество и запасенное в ней энергия производят сами сообщества называется автохтонным, то, что поступает в экосистему из вне - аллохтонным. Соотношение автохтонного и аллохтонного вещества и энергии изменяется в экосистемах в пространстве и во времени.
Экологическая эффективность. Значительный интерес представляет соотношение величин энергетического потока в разных точках пищевой цепи выраженных в %. Эти отношения называются экологической эффективностью. Основных показателей три:
-
Эффективность потребления ЭП = 100 Pn/In -1
ЭП - коэффициент суммарной продукции первого трофического уровня, которое потребляется следующим трофическим уровнем остальное живое существо гибнет самостоятельно и поставляется в систему редуцентов. Среднее значение эффективности потребления для лесов около 5%., для степей 20-25%, для фитопланктона около 50%.
-
Эффективность ассимиляции ЭА = 100 An/In1, т. е. процесс энергии пищи попадает в пищеварительную систему консумента не некотором трофическом уровне. Эта энергия становиться доступной в ходе роста или совершения работы. Остальное теряется с выделением и поступает в систему редуцентов. Для микрофилов и грибов это соотношение не подходит.
Эффективность ассимиляции низкая у фитофагов, микробофагов, дитритофагов = 20-50%, у хищников достигает 80%, у фитофагов в кишечнике собственная система редуцентов.
-
Эффективность продуцирования ЭПр= 100 Pn/An
Это процент ассимилированной энергии включаемой в биомассу нового трофического уровня. Остальное теряется в процессе дыхания и с продуктами выделения. Эффективность продуцирования изменяется в зависимости от систематического положения организма. У беспозвоночных 30-40%, у позвоночных около 10%, у эндотермов около 1-2%.
-
Эффективность использования пищи на рост. В 1983 г Ивлев ввел два трофических коэффициента, которые характеризуют эффективность использования пищи на рост.
К1 = Q1 : Q K2 = Q1 : (Q-Q2)
К1 - определяется по первому уровню.
Q1 = энергии прироста, т. е. энергия вновь образуемого в организме вещества.
Q - энергия потребляемой пищи.
К2 - трофический коэффициент второго порядка.
Q2 - энергия не усвоенной пищи.
Так как, (Q - Q2) - энергия ассимилированной пищи, которая расходуется на прирост тела (Р) и энергозатраты (Т).
К2 = Р : (Р+Т) - Вимперг
Эта функция позволяет судить о КПД организмов и популяций в том случае, если известны их прирост и энергетические затраты.
Величина трофического коэффициента видоспецифична и зависит от состава популяции и внешних условий. С возрастом К2 снижается достигая 0 при приближении к полному размеру тела. У голодных животных используемых пищу на рост ниже чем у сытых. С изменением условий питания, когда на добывание пищи тратиться меньше энергии К2 - возрастает. К2 резко снижается с ухудшением условий дыхания и с повышением температуры или когда популяция работает в режиме отклоняющимся от оптимального. Одной из характеристик использования пищи является степень ее усвоения, т. е. переваривания, следовательно, это соотношение ассимиляции и потребления. У растительноядных форм этот показатель значительно ниже, чем у плодоядных. Степь ассимиляции потребленной пищи сильно меняется в зависимости от ее качества, физиологических особенностей потребителя и условий питания. Взаимосвязь между величиной потребляемой энергии, т. е. рационом (Р), степенью усвоения пищи (У), приростом (П) и тратами на обмен (Т) выражается балансовым равенством
УП = П+ Т
Р = У (П+Т).
При определении величины рациона важно учитывать те питательные вещества, которые воспринимаются астматически могут играть весьма существенную роль в энергообеспечении гидробионтов.
Роль консументов в динамике пищевой сети.
Животные и другие консументы - это не пассивные «едоки», входящие в пищевую цепь. Удовлетворяя свои потребности в энергии, консументы часто действуют через систему положительной обратной связи на находящимся выше трофическом уровне. Благодаря естественному отбору хищники и паразиты приспособились к тому, чтобы не только не уничтожать источники своей пищи, но во многих случаях обеспечивать или даже увеличивать благосостояние своих жертв. Так что теоретически должны существовать не только управляющие цепи отрицательной обратной связи, но и эффекты положительной связи.
Некоторый интерес представляет длина пищевой цепи. Однако доступность энергии, видимо не единственный фактор, поскольку длинные пищевые цепи часто встречаются в неплодородных системах, например в олиготрофных озерах, а короткие - в продуктивных или эвтрофных системах. Быстрое продуцирование питательного растительного материала может стимулировать быстрое выедание в результате чего поток энергии концентрируется на первых двух - трех трофических уровнях.
Кроме консументов в регуляции использования первичной продукции участвуют и другие механизмы. Это, например, химические соединения, выедаемые, выедаемые растениями и ингибирующие гетеротрофное потребление.
В 1960 году Хэйрстон, Смит и Слободки сформулировали гипотезу «естественного равновесия». Авторы гипотезы предположили, что растения в общем накапливают очень большую биомассу, видимо что-то препятствует их выеданию это по их мнению делают хищники, т. е. численность первичных консументов ограничивается вторичными консументами, а первичные продуценты, таким образом, ограниченны не выеданием травоядными, а только ресурсно.
Концентрация токсичных соединений при продвижении по пищевым цепям.
Распределение энергии, разумеется не единственный количественный параметр, на который влияют события происходящие в пищевых цепях. Некоторые вещества по мере продвижения по цепи не рассеиваются, а наоборот, накапливаются. Это так называемое концентрирование в пищевой цепи (биологическое накопление).
Тенденции некоторых радионуклидов, побочных продуктов деления ядра атома, увеличивать свою концентрацию с каждым этапом пищевой цепи впервые была обнаружена на Хелфордском заводе. Крайне малые количества радиоактивного йода, фосфора, цезия и стронция выпускавшегося заводом в реку Колумбии, как оказалось, концентрировалось в тканях рыб и птиц. Изучая модель детритной пищевой цепи, можно понять, что благодаря многократному поглощению с начала детритной пищевой цепи в ней должно накапливаться любое вещество быстро собирающееся на частицах детрита и почвы и растворяющееся в кишечнике.
Качество энергии.
Энергия характеризуется не только не только количественным, но качественным параметром. Не все калории одинаковы, т. е. одинаковые количества разных форм энергии могут сильно различаться по своему рабочему потенциалу. Высококонцентрированные формы, такие как энергия нефти, обладают более высоким потенциалом и соответственно более высоким качеством, чем такие «разбавленные» формы, как солнечный свет, а солнечный свет в свою очередь обладает более высоким качеством по сравнению с еще более рассеянным низкотемпературным теплом. Качество энергии измеряется ее количество, расходуемым при превращении, или говоря точнее, количеством определенного типа энергии, затрачиваемого на получение другого типа в цепи превращений энергии. По мере того как в этой цепи уменьшается количество энергии, пропорционально на каждом этапе повышается количество той ее доли, которая действительно перешла в новую форму.
В сущности, с точки зрения термодинамики качество энергии измеряется расстоянием, пройденным его от солнца. Произведение действий потока энергии на каком - то уровне на показатель качества называется затраченной этого компонента. Доступна ли консументу потонциальная энергия, содержащаяся в компоненте, зависит от качества ресурсов, например его пищевой ценности.
Качество энергии небольшой биомассы хищников в 100 раз выше, чем такой же биомассы травоядных животных. Это более высокое качество отражается в управляющем воздействии компонентов, лежащих «ниже по течению» энергии, на компоненты, лежащие выше.
Общая геохимическая организация биосферы.
Химический состав земной коры, как фактор биосферы.
Земные наружные оболочки земли, различающиеся фазовым состоянием: твердая - земная кора, жидкая - гидросфера и газовая - атмосфера. Тесно связаны между собой, а вещество каждой из них проникает в пределы других, например, подземные воды, проникают в верхнюю часть земной коры, значительный объем газов находиться не в атмосфере, а растворен в гидросфере и заполняет пустоты почвы в горных породах. В свою очередь вода и мелкие твердые минеральные частицы насыщают нижние слои атмосферы следовательно, в земной коре основной резерв химических элементов, которые вовлекаются в миграционные процессы под воздействием живого вещества.
Относительное содержание химических элементов в земной коре.
Химики начиная со второй половины 19 века изучали химический состав горных пород методами весового и объемного химического анализа. Суммируя результаты многочисленных анализов горных пород Кларк показал, что в земной коре преобладаю кислород, кремний, алюминий, железо, магний, кальций, калий и натрий. Появился новый метод спектораскопического анализа. Новые факты позволили Вернадскому сформулировать принцип «всюдности» всех химических элементов. Идея «всюдности» химических элементов долгое время вызывала настороженность. В итоге усилий ученых разных стран в 20-х годах 20 века сложилось общее представление о составе земной коры. Среднее значение относительного содержания химических элементов в земной коре и других глобальных и космических системах известных геохимик Ферсман предложил назвать Кларками. Анализ значений Кларков позволяет понять многие закономерности распределения химических элементов на земле, в солнечной системе и доступной нами наблюдаемой части Вселенной. Кларки химических элементов земной коры различаются более, чем на десять математических порядков. Столь существенное количественное различие должно отразиться на качественно неодинаковой роли двух групп элементов в земной коре. Элементы первой группы называются главными, второй - рассеянными.
Формы нахождения химических элементов в земной коре.
Не смотря на то, что общее количество природных химических соединений - минералов составляют 2 - 3 тысячи видов числа минералов образующих распространенные горные породы, невелико. Более 80% массы земной коры представлено силикатами алюминия, железа, кальция, магния, калия и натрия. Около 12% составляет оксид кремния. Все эти минералы имеют кристаллическое строение, которое определяет общие особенности кристалохимии земной коры. Согласно концепции Гольдшмидта в кристаллохимических структурах ионы ведут себя, как жесткие сферы твердого шара. Поэтому радиус каждого иона рассматривается как постоянная величина. Главная особенность ионов в кристаллохимических структурах заключается в том, что радиусы отрицательно заряженных ионов (анионов) значительно больше радиуса положительно зараженных ионов (катионов).
Образование распространенных минералов сопровождается своего рода сортировкой рассеянных элементов.
В распространение минералов и горных пород избирательно задерживаются химические элементы концентрации, которых не так велика для образования самостоятельных минералов. Взаимное замещение ионов в кристаллической структуре благодаря близости их радиусов называется изоморфизмом. В результате изоморфизма рассеянные элементы закономерно концентрируются в определенных минералах. Полевые шпаты служат носителями бария, стронция, свинца, олова - никеля и кобальта, цирконы гафния .
Изоморфное замещение не единственная форма нахождения рассеянных элементов. рассеяния в земной коре проявляется в разных формах на разных уровнях дисперсии.
Применение поляризованного микроскопа и прозрачных шлифов позволило обнаружить в минералах не только твердые включения и микропустоты заполняемые остатками растворов из которых осуществлялась кристаллизация. Это явление специально рассматривалось в 1858 году основателем оптической петрографией Г. Сорби в настоящее время всесторонне изучено.
Результаты многих технологических анализов выполненных в разных странах за последние 50 лет позволяют утверждать, что все парообразные минералы являются носителями рассеянных элементов. Именно в них сосредоточенна основная масса рассеянных элементов, содержащихся в земной коре. Зная содержание минералов - носителей и концентрация в них рассеянных элементов можно рассчитать баланс внутри конкретной горной породы.
Формы нахождения рассеянных элементов в кристаллическом веществе земной коры.
-
Микроминералогические формы:
- элементы, входящие в акцессорные минералы;
- элементы, содержащиеся в микроскопических выделениях в результате распада твердых растворов;
- элементы, находящиеся во включениях остаточных растворов.
2. Неминералогические формы:
- элементы сорбированные поверхностью дефекта реальных кристаллов;
- элементы, входящие в структуру минерала - носителя по законам изоморфизма;
- элементы, находящиеся в структуре минерала - носителя в неупорядоченном состоянии.
Сочетание рассмотренных форм нахождения рассеянных элементов сильно меняется в зависимости от многих факторов. Соответственно и меняется суммарное содержание рассеянного элемента в различных участках земной коры. Поэтому для объективной оценки содержания элемента используют методы математической статистики.
Чтобы количественно оценить неоднородность химических элементов в земной коре Вернадский ввел специальный показатель - Кларк концентрации.
Кк = А/К
Его числовое значение характеризует отклонение содержание элемента в данном объеме от Кларка:
А - содержание химических элементов в горной породе, руде и минерале и др.
К - Кларк этого элемента в земной коре.
Если Кк > 1 это указывает на обогащение элементом.
Если Кк < 1 это означает снижение его содержания по сравнению с данными земной коры в целом.
Изменение концентрации химических элементов в пространстве, отклонение от глобальной или местной геохимической нормы, это не отдельные случаи, а характеризуется черта геохимической структуры земной коры. Это имеет очень важное значение для состава фотосинтезирующих организмов суши, которые образуют основную часть массы живого вещества земли.
Биогеохимия газовой оболочки Земли.
Становление Земли как планеты сопровождалось формированием его зонального строения и общепланетарной дифференциацией химических элементов. Принято считать, что первым результатом дифференциации было образование тяжелого ядра и силикатной мантии, результаты изотопного анализа дают основание предполагать, что возникновение наружных фазовых оболочек твердой, жидкой и газовой так же произошло на раннем этапе развития планеты.
С момента обособления началась эволюция их состава, в которой выдающуюся роль сыграло живое вещество. Наиболее сильное воздействие биогеохимических процессов сказалось на составе атмосферы.
Биогеохимическая эволюция состава атмосферы и жизнедеятельность организмов в массообмене газов.
История формирования атмосферы служит ярким примером воздействия живого вещества на окружающую среду. Полученные в последние годы факты свидетельствуют, что состав современной газовой оболочки земли является итогами длительно процесса, в котором ведущее значение имеет геохимическая деятельность живых организмов.
Масса атмосферы составляет около 5,2*10 в 15 степени тонн. Основная часть газового вещества (80%) заключена в тропосфере. Верхняя граница которой расположена на высоте около 17 км на экваторе, к полюсам она снижается до 8-10 км.
Верхняя граница тропосферы - тропопауза. Она начинает область сильного снижения температуры и отсутствия скопления паров воды. Тропосфера является областью активного взаимодействия с океаном и с сушей, в ней сосредоточена основная масса паров, воды и мелких твердых частиц переносимых воздушными массами. В тропосфере происходит фотохимические реакции имеющие важное значение для биосферы.
Выше тропопаузы в стратосфере и мезосфере нарастает разреженность газов, сложно меняются термические условия. На высоте 25-30 км под воздействием солнечной радиации происходит фотодиссоциация молекул кислорода и образуется озон. Молекулы озона сильно рассеяны. Если бы они находились в условии обычного для поверхности земли мощность озонового слоя была бы менее 4 см. очень разреженный слой озона поглощает 97% ультрафиолетовых частиц солнечной радиации. Без этого экрана существование жизни на поверхности суши была бы невозможной. На удалении от 80-800 км от поверхности земли располагается ионосфера - область сильно разреженного ионизированного газа. Самая наружная часть газовой оболочки - это экзосфера. Она простирается до 1 800 км. Из этой сферы происходит диссипация - это потеря землей наиболее легких атомов водорода и гелия. Состав газовой оболочки земли, как и состав океана в значительной мере обусловлен деятельностью живых организмов и поддерживается системой биогеохимических циклов. В настоящее время газовое вещество атмосферы на 99,9 % состоит из озона, кислорода и аргона среди компонентов содержащихся в малых количествах можно выделить пары воды, инертные газы и соединения, обусловленные биологическими процессами и фотохимическими реакциями.
Большой интерес для реконструкции истории атмосферы представляет геохимия инертных газов. Относительное высокое содержание аргона связано с тем, что большая часть этого газа представлена изотопом аргона, образованного за счет распада радиоактивного изотопа калия. Количество гелия в атмосфере наоборот в тысячи раз меньше, чем должно быть. Это обусловлено непрерывной диссипацией этого элемента. Остальные инертные газы содержатся в том количестве в каком они были выделены на протяжении всего времени существования Земли.
Изучение соотношения изотопов ксенона привело геохимика Ю. А. Шуколюкова к заключению, что газовая оболочка возникла за очень короткий отрезок времени, который совпадает со временем акреции (сгущение, агрегация) земли. Предполагают, что быстрое образование атмосферы обусловлено энергичным выделением газов при ударах метеоритных тел на ранней стадии развития земной коры.
Под воздействием солнечной радиации в атмосфере происходила диссоциация паров воды. Непрерывное удаление глобального восстановителя (водорода) в результате диссипации вызывало прогрессирующее накопление окислителя. Это имело два важных последствия: 1. постепенно образовывался экран предохраняющий поверхность земли от ультрафиолетовой радиации; 2. возник процесс окисления, восстановленных газов непрерывно поступающих из мантии.
Окислители фотохимического происхождения постоянно возобновлялись в результате диссипации водорода. Окисленные соединения растворялись в конденсирующейся атмосферной влаги и вымывались из атмосферы дождями. Этот процесс, имевший циклический характер имел направленность будущих биогеохимических циклов.
Хемолитотрофные сообщества включавшиеся в механизм воздействия окислителя атмосферы с восстановленными газами выделявшимися из недр земли знаменуют первый этап геохимической деятельности живого вещества.
Вхождение организмов в глобальную систему миграции, выделения газов из земли в атмосферу и окисление и последующее вымывание из атмосферы. Это усложнило систему и превратило ее из абиогенной в биогенно-абиогенную.
Следующий этап связан с распространением цианобактерий (сине-зеленых водорослей), которые для синтеза органического вещества начали использовать не энергию окислительно - восстановительных химических реакций, а световую энергию солнца. Признаки деятельности этих простых форм жизни отмечены уже в самых древних геологических образованиях. В кремнистых образованиях, возраст которых определяется в 3,5 млрд лет уже обнаружены строматолиты - это структуры созданные цианобактериями. Возможно присутствие свободного кислорода было не повсеместно, а лишь на отдельных участках. При реакции фотосинтеза кислород стал выделяться в качестве метаболита. Для цианобактерий свободный кислород токсичен, при его отсутствии они нормально развиваются.
Около 1,5 млрд лет назад произошел переход от цианобактериальных сообществ к сообществам альгобактериальным. Вероятно определенную роль в этом событии сыграло накопление свободного кислорода в океане и атмосфере и как результат - создание новых условий, в которых конкурентность цианобактерий была не высокой . в глобальном процессе создания органического вещества водоросли постепенно заместили цианобактерии. В конце протерозоя на протяжении венда сложилась система из продуцентов - фотосинтетиков и консументов - животных, обуславливающая углерод-кислородный биогеохимический цикл.
Формирование химического состава атмосферы происходило путем закономерной дифференциации химических элементов выделяемых из недр Земли в виде восстановленных газов. Система обеспечивающая указанную дифференциацию, изначально абиогенная, и имевшая циклический характер являлось главной и характерной чертой среды развития древнейших организмов, которые заняли в ней определенное место. В начале биохимические процессы выполняли роль отдельных звеньев в системе глобального круговорота газов в атмосфере. В дальнейшем по мере развития форм жизни циклическая миграция газов. Стала контролироваться жизнедеятельность организмов. Это оказывало влияние на состав атмосферы, океана и горных пород на поверхность древней суши. В свою очередь, изменение Эколого-геохимических условий среды обитания оказало воздействие на развитие организмов и совершенствование структуры биогеохимических циклов. Со времени широкого распространения эукариот ведущим стал углерод, кислородный цикл.
Биокосная система гидросферы.
Вода имеет большое значение для живого вещества земли. Она является обязательным условием его существования и главной составной частью.
Современный химический состав океана представляет собой черты его длительных изменений под действием жизнедеятельности живых организмов.
Среди продуктов дегазации преобладают пары воды и углекислый газ. С того момента как температура поверхности планеты опустилась ниже 100ºС пары воды стали конденсироваться не образовывая первичные водоемы. На поверхности земли возник круговорот воды.
Появление цианобактерий обитавших в воде, предохранявшей их от губительной ультрафиолетовой радиации, ознаменовало начало фотосинтеза и биогеохимического продуцирования кислорода.
Свободных кислород стал поступать в воды древнего океана. Состав океанической воды приобрел хлоридно - сульфатный состав близкий к современному.
Химические элементы в гидросфере находился в разных формах. Особую группу составляют элементы органических соединений.
Ученые обнаружили геохимическую особенность океанической воды. Соотношение главных ионов остается постоянным. Солевой состав океана является своего рода геохимической константой. Ведущим фактором эволюции химического состава океана на протяжении геологической истории была суммарная биогеохимическая деятельность живых организмов.
Среди пелагических илов (глубинные отложения океана) можно выделить две группы: 1. состоит из биогенных образований планктонов; 2. образованно в основном частицами не биогеохимического происхождения. В первой области наиболее распространены известковые (карбонатные илы). Во второй - глинистые илы.
Ежедневное накопление высокодисперсного органического дитритуса в осадках мирового океана превышает 0,5 * 10 в девятой степени тон. Дисперсное органическое вещество сорбируют увлекая в осадок комплекс рассеянных элементов. В органическом веществе активно накапливаются многие токсичные элементы (мышьяк и свинец), которые удаляют из океанической воды дисперсное органическое вещество. Взвеси выполняют роль глобально сорбента регулирующего содержание рассеянных элементов и предохраняют среду мирового океана от опасных уровней и концентраций.
Особенности геохимии поверхности вод суши.
Океана пополняется за счет стока воды с суши, который примерно равен 44 * 10 в третей степени кубических метров в год. Динамическое постоянство объема океана поддерживается испарением и переносом через атмосферу в парообразном состоянии в таком же количестве воды.
Речные воды создают мощный геохимический поток, играющий важную роль в общепланетарном массообмене между мировой сушей и океаном. В речных водоемах различают следующие главные формы химических элементов: 1. простые и комплексные ионы; 2. нейтральные молекулы; 3. коллоидные частицы состоящие из устойчивых гумусовых веществ, аморфных процессов разрушения глинистых минералов оксида железа; 4. крупные взвешенные частицы.
Соотношение масс растворенных соединений твердых взвесей в речном стоке в значительной мере зависит от характера растений суши.
В речных водах содержаться растворенные формы рассеянных элементов, не захваченных в биологический круговорот. В тоже время, находящееся в растворе элементы могут образовывать комплексные соединения с органическими веществами, сорбироваться гелием в макромолекулы, а высокодисперсные частицы - илы и осадки, в таких условиях, когда теоретически этого не должно быть. Значительная часть рассеянных элементов присутствует в природных водах в форме комплексных соединений. Важное значение для водной миграции имеют комплексные органические, особенно внутрикомплексные соединения металлов. В этих соединениях ионы металлов имеют ионную и координационную связь с отдельными функциональными группами внутри молекулы. Несмотря на то, что общая минерализация вод значительно меньше соленых морских, глобальный вынос рассеянных элементов в растворах весьма значительный. Отдельные элементы выносятся в меньших количествах.
Интенсивность водной миграции химических элементов.
Преставление о выносе растворенных масс элементов должно быть должно быть дополнено оценкой степенью интенсивности вовлечений их в водную миграцию.
В 1874 г. ученые предложили коэффициент водной миграции (Кв), который равен отношению концентраций элементов в сухом остатке воды и в породе, оценивая интенсивность вовлечения элемента водной миграции.
Эволюция биосферы.
Еще недавно, эволюция рассматривалась с точки зрения формирования биологических систем. Самый длинный отрезок в истории земли приходился на образование биосистем из неорганической материи. Несколько меньше времени понадобилось на возникновение первых клеток и лишь после этого начался все ускоряющийся процесс собственно биологической эволюции. Первыми живыми существами были гетеротрофные микроорганизмы питавшиеся «первичным бульоном», той органикой, которая в избытке возникала в первичной атмосфере и океане, в результате процессов частично смоделированных Миллером.
За тем уже возникли автотрофы синтезирующие органику из углекислого газа и воды, используя для этого энергию окислительных химических реакций или солнечного света. Реальные факты вынудили отказаться от этой умозрительной схемы, не нашли подтверждений об исходной гетеротрофности живых организмов. Судя по всему авто и гетеротрофы возникли практически одновременно. Один из ведущих специалистов по микробиологии Заварзин обращает внимание на то, что «первичный бульон» даже теоретически не может быть источником для первичной жизни, так как он является исчерпаемым ресурсом. Поскольку живые существа размножатся в геометрической прогрессии, потомство первых гетеротрофов должно было съесть этот бульон за короткий промежуток времени.
Ни автотрофная ни гетеротрофная теории не могли были быть. Первичность хемоавтотрофности тоже было принято считать несомненным (аргументом является то обстоятельство, что наиболее древние из прокариот археобактерий).
Хлорофилл поглощает квант света, в дальнейшем энергия этого кванта используется в различных химических превращениях, сейчас есть основание предполагать, что первичная функция этих пигментов была нейтрализация разрушительных энергетических квантов ультрафиолетового излучения. В последствии те организмы, которые приспособились, еще и использовать эту энергию «в мирных целях» получили гигантские преимущества. Однако сама по себе «радиационная» защита ДНК при помощи пигментов должна была сформироваться еще на стадии доорганизменных гиперциклов.
Понятно, что доводить эту защиту до совершенства имеет смысл лишь тем организмам которые обитают в поверхностном слое океана (10 м - защита от ультрафиолета). Именно эти организмы дали начало фотоавтотрофам, тогда как глубины остались в распоряжении гетеротрофов.
Существование фотоавтотрофов с самых древних времен есть прямые доказательства. Древние организмы из формации Исуа были именно синтезирующими, а в породах возраста около 3 миллиардов лет содержаться остатки хлорофилла (фитан, фикабелин - пигмент), но гораздо проще отслеживать деятельность фотоавтотрофов по одному из ее побочных продуктов (по кислороду).
Впервые такой анализ был проведен в 60-е годы Бекнером и Маршаллом. Они пришли к выводу, что количество кислорода на Земле росло на протяжении всей её истории, постоянно, но крайне медленно. Кислород - не появился вследствие дегазации магмы, а следовательно полностью отсутствовал в первичной атмосфере Земли. Об этом свидетельствует наличие в древних, главным образом катархейских породах легко окисляемых, но не окисленных веществ (графит, лазурит и т.п.). Небольшие количества кислорода образуются в результате фотодиссоциации молекул воды под воздействием жесткого ультрафиолетового излучения, полностью расходовались на окисление постоянно выделяемых вулканами аммиака, метана, сероводорода. Однако, начиная с определенного момента, в океанах начинали образовываться джеспилиты (полосчатые железные руды), представляют собой чередование бедных и обогащенных окислами железа слоев. Они формируются когда двух валентное (закислое) железо сформировавшиеся в результате вулканической активности и растворено в морской воде реагирует в поверхностных слоях океана с молекулами кислорода, окисляется до трехвалентного (окислого) и выпадает в виде нерастворенного осадка на дно. Таким образом начала образовываться руда этого типа (состоит из гематита Fe2O3 и магнетита FeО ∙ (Fe2O3)), означает появление на Земле достаточно мощного источника молекул кислорода, которым могут быть только фотосинтезирующие организмы. Впервые джестилиты появились в Исуа, а это может служить подтверждением того, что содержимое в этой формации углеродистое вещество действительно принадлежит фотосинтетикам.
Источник кислорода возник, но мир еще в течении 1,5 млрд лет оставался анаэробным. Об этом свидетельствует наличие в отложениях конгломератов из спирита (FeS2) и ряда других минералов являющихся термодинамично не устойчивыми в присутствии кислорода. Зачастую они представляю собой гальку с отчетливыми следами обработки ее течением - это первое свидетельство существования на Земле пресных вод. Но текучие воды не могут не быть хорошо аэрированными, и то, что пирит оставался неокисленным свидетельствует о практически безкислородной атмосфере.
Сообщество фотоавтотрофов формируют в это время своеобразные кислородные оазисы (Термин ввел Клауда). Их возможностей хватало на создание окислительной обстановки и осаждение железа в окисленной форме, только в своем непосредственном окружении. Ситуация радикально изменилась около 2 млрд лет назад, когда процесс гравитационной дифференциации недр привел к тому, что большая часть железа перешла в ядро планеты и растворенное в морской воде закислое железо не могло уже возобновляться по мере выбывания из цикла (биогеохимический цикл железа). Фотоавтотрофы получили возможность завершить свою работу по окислению закислого железа и осаждению его в виде джестилитов. В это время возникли все крупнейшие месторождения железа. В дальнейшем руды этого типа не образовывались.
Кислород ранее полностью расходовался на этот процесс, теперь стал насыщать атмосферу. С этого времени пиритовые конгломераты исчезли, а на смену донно-морским железным рудам прийти терригенные красноцветы. Яркий образ для описания этих событий придумал Заварзин. Он назвал их временем, когда биосфера вывернулась наизнанку за счет появления кислородной атмосферы.
Именно в это время 1,9 млн лет назад в Канадской формации Ганфлинд впервые появились извесчатые основания полностью идентичные тем, которые образуют в настоящее время облигатно аэробное марганцевое- осаждающая бактерия Metallogenium. Без кислорода окисление железа и марганца не идет, а образуемые этой бактерии ей металлические в виде характерных «паучков» возник только в сильно окислительной обстановке, следовательно это должно означать, что в этот момент содержание кислорода в атмосфере уже достигло величины, как 1% от современного (точка Пастера). Именно с этой пороговой концентрации становиться экономично оправданным кислородное дыхание, в ходе которого из каждой молекулы глюкозы можно будет получать 38 молекул АТФ, вместо 2 при кислородном брожении. С другой стороны, в атмосфере начинает возникать озоновый слой, преграждающий путь смертоносному ультрафиолету, что ведет к колоссальному расширению спектра пригодных для жизни местообитаний. Примерно к середине протерозоя (1,7-1,8 млрд лет назад) "кислородная революция" в целом завершается и Мир становится аэробным. Впрочем, с точки зрения существ, составлявших тогдашнюю биосферу, этот процесс следовало бы назвать иначе: "Необратимое отравление кислородом атмосферы планеты". Смена анаэробных условий на аэробные не могла не вызвать катастрофических перемен в структуре тогдашних экосистем, и в действительности "кислородная революция" есть не что иное, как первый в истории Земли глобальный экологический кризис.
Итак, в раннем докембрии существовал совершенно особый мир, формируемый прокариотными организмами - бактериями и цианобактериями. Разделение живых существ на прокариоты и эукариоты (эти термины были введены в 1925 г. Э.Шаттоном), основанное на наличии или отсутствии в их клетках оформленного ядра, теперь считают существенно более фундаментальным, чем, например, разделение на "животные" и "растения". Среди многих их различий для нас сейчас важнее всего то, что среди прокариот неизвестны многоклеточные организмы. Есть нитчатые и пальмеллоидные формы цианобактерий, однако уровень интеграции клеток в этих структурах - это все-таки уровень колонии, а не организма. А поскольку времени на всякого рода эксперименты у прокариот, как мы видели, было более чем достаточно (как-никак, три с половиной миллиарда лет), то приходится предположить, что по каким-то причинам на прокариотной основе многоклеточность не возникает в принципе. Может быть дело в отсутствии у них центриолей и митотического веретена, без которого невозможно точно ориентировать в пространстве делящиеся клетки и формировать из них сколь-нибудь сложные ансамбли. Может быть - в свойствах их клеточной оболочки, препятствующих межклеточным взаимодействиям; это делает невозможным обмен веществами, так что клетки, находящиеся внутри организма и не имеющие прямого контакта с окружающей средой (что неизбежно при многоклеточности), не могли бы питаться и выводить продукты распада. Может быть - в чем-то еще; как бы то ни было, такой путь повышения уровня организации, как многоклеточность, для прокариот оказался закрытым. Выход из этого тупика они нашли весьма оригинальный.
Для докембрийских осадочных толщ чрезвычайно характерны строматолиты (по-гречески - "каменный ковер"). Это тонкослойчатые колонны или холмики, состоящие главным образом из карбоната кальция. Выяснилось, что строматолит образуется в результате жизнедеятельности совершенно ни на что не похожего прокариотного сообщества, называемого цианобактериальным матом. Мат, располагающийся на верхней поверхности создаваемого им строматолита, представляет собой плотный многослойный "ковер" общей толщиной до 2 см; основу его составляют нитчатые либо пальмеллоидные цианобактерии, однако помимо них в формировании сообщества участвует множество самых различных микроорганизмов. Внутри мата легко различимы несколько функционально дискретных слоев:
1. Плотный верхний слой - поверхность роста (1 - 1,5 мм), в котором есть и автотрофы, продуцирующие кислород цианобактерии-фотосинтетики, и гетеротрофы - облигатно-аэробные (неспособные жить в отсутствии кислорода) бактерии.
2. Тонкая (менее 1 мм) подкладка; в ней автотрофами служат бактерии, осуществляющие нескислородный фотосинтез (при котором источником водорода является не вода, а другие вещества, например, сероводород - тогда выделяется не молекулярный кислород, а сера). Гетеротрофами же служат факультативные аэробы, которые могут использовать кислород, но в его отсутствие способны удовлетворять свои потребности в энергии не дыханием, а брожением.
3. Ниже двух верхних слоев, в которых идут процессы фотосинтеза и которые в совокупности называют фотической зоной мата, лежит толстая бескислородная зона, в которой процветают разнообразные анаэробы.
На поверхность обитающего на мелководье мата постоянно выпадают частички осадка (обычно - кристаллы карбоната кальция), которые затемняют соответствующе участки фотической зоны и должны постепенно ухудшать условия происходящего в ней фотосинтеза - вплоть до неизбежной гибели фотосинтетиков. От катастрофы эти микорорганизмы спасает положительный фототаксис: по мере того, как интенсивность света в фотической зоне падает, они мигрируют вверх сквозь накапливающийся слой осадка, образуя выше него новую поверхность роста с подкладкой. Миграция эта идет разными способами: нитчатые формы обладают способностью к скользящему движению сквозь осадок, пальмеллоидные - к ускоренному нарастанию верхних частей колонии; еще один путь - временный переход в латентное состояние с последующей реколонизацией поверхности осадка.
Не все маты представляли собою донные сообщества. Дело в том, что, помимо уже известных нам строматолитов, следы жизнедеятельности микробных сообществ представлены еще и онколитами - более или менее сферическими образованиями, в которых известковые слои располагаются не линейно (как в строматолите), а образуют концентрическую структуру.
Итак, мат представляет собой высокоинтегрированное сообщество с чрезвычайно сложно организованной трофической структурой: два верхних слоя составлены продуцентами и консументами первого порядка ("растениями" и "травоядными"), соединенными в пищевую цепь пастбищного типа, а нижний слой сформирован редуцентами ("падальщиками"), получающими органику из верхних слоев по детритной цепи. Более того, измерения показали, что мат является вообще одной из самых сбалансированных экосистем: он производит ровно столько органики и кислорода, сколько тут же расходует в процессе своей жизнедеятельности (нулевой баланс); а ведь замкнутость геохимических циклов была ранее определена нами как главное направление в эволюции биосферы. Именно такая структура "производства и протребления" в прошлом позволяла матам образовывать кислородные оазисы и постепенно увеличивать их - вместо того, чтобы пытаться сходу изменить всю эту бескислородную среду.
Можно сказать, что в мате "ничто не пропадает" - даже энергия солнечного света. Фотосинтезирующие бактерии подкладки имеют пигменты, позволяющие им поглощать свет из иной части спектра, чем их "коллеги"-цианобактерии из вышележащего слоя. Таким образом, поверхность роста становится для жителей подкладки как бы прозрачной, что позволяет утилизовать всю поступающую в фотическую зону световую энергию. Более того, поскольку солнечный спектр меняется в течение дня ("покраснение" Солнца на восходе и закате), для улучшения использования энергии в мате происходят упорядоченные вертикальные миграции - смена горизонтов, занимаемых бактериями с разными типами пигментов.
Все это позволяет сделать следующий вывод: уровень интеграции, достигнутый составляющими мат микроорганизмами, превосходит уровень, наблюдаемый в обычных экосистемах, и как минимум не уступает тому, что наблюдается у лишайников. И уж коль скоро мы считаем "организмами" лишайники, то такое определение с полным основанием можно отнести и к мату.