- Преподавателю
- Биология
- Урок биологии 5 класс
Урок биологии 5 класс
| Раздел | Биология |
| Класс | 5 класс |
| Тип | Конспекты |
| Автор | Феоктистова Т.А. |
| Дата | 04.02.2015 |
| Формат | docx |
| Изображения | Есть |

Проведённый урок является пятым в разделе «Ткани живых организмов» курса биологии в 5 классе. Следующий урок - обобщающий по данному разделу.
Учебное занятие разработано в соответствии с программой основного общего образования по биологии под редакцией Сухоруковой Л.Н.Тема урока соответствует рабочей программе и тематическому планированию по предмету.
Специфика урока заключается в том, что этот урок-исследование, где выполняется лабораторная работа «Рассматривание под микроскопом готовых микропрепаратовмышечных и нервной ткани животных» и используются ЭОР. Использование электронного учебника позволяет за ограниченное рамками урока время дать больший по объему материал, иллюстрирующий нужную информацию.
Тип урока - учебное занятие по изучению и первичному закреплению новых знаний и способов деятельности. На занятии продемонстрирован системно-деятельностный подход, когда усвоение происходит только через собственную деятельность.
Выбор структуры урока обусловлен тем, что каждый следующий этап урока является логическим продолжением предыдущего. На уроке задан достаточно высокий темп в силу объемности материала. Выбор содержания данного урока обусловлен тем, что через него решаются цели и задачи, поставленные вначале урока.
На уроке преобладает частично-поисковый характер деятельности учащихся, постановка проблем, решения которых лежат за пределами изучаемого курса, создаются условия для освоения учащимися знаний преимущественно в форме деятельности.
Высокая работоспособность учащихся на уроке достигается за счет познавательного интереса, за счет навыков выполнения практических работ. Урок непосредственно связан с предыдущими уроками в данном разделе, так как опирается на знания учащихся о строении клетки и её жизнедеятельности, строении тканей животных и растений.
Данный урок является многоцелевым, поскольку позволяет решить комплекс задач:
- изучить новый материал о мышечных и нервнойтканях;
- прививать навыки самостоятельного поиска и навыки работы в сотрудничестве;
- пробуждать любознательность;
- воспитывать вежливое, уважительное отношение друг к другу, умение вместе обучаться.
Были соблюдены основные принципы обучения: последовательность, доступность, наглядность, активность, научность.
В целом, удалось решить на необходимом уровне поставленные задачи урока и получить соответствующие им результаты обучения, избежав при этом перегрузки и переутомления учащихся, сохранив рабочее настроение и хорошее самочувствие.
Технологическая карта урока
Предмет
биология
Класс
5
Тема урока
Мышечные и нервная ткань животных
Тип урока
Учебное занятие по изучению и первичному закреплению новых знаний и способов деятельности
Цели
Создать условия для:
- систематизации знаний учащихся о строении и жизнедеятельности тканей животных;
- формирования представлений о ткани, клетке как биологической системе (целое, состоящее из частей, выполняющих определенные функции);
- формирования представлений у учащихся о тканях животных, их видах, особенностях строения и функций, а так же о месторасположении различных тканей в организме животных и человека.
- формирования умений работать с рисунками, тренажёрами, микроскопами;
- развития воображения, мыслительных операций: анализ, синтез, сравнение, обобщение
- воспитания мотивов, направленных на изучение живой природы и бережного отношения к ней;
Оборудование:
компьютер, мультимедийный проектор, интерактивная доска
микроскопы, микропрепараты тканей животных, гистологические микрофотографии.
Формы работы:
индивидуальная, парная, групповая, фронтальная.
Методы проведения урока:
практический метод - лабораторная работа учащихся, самостоятельная работа с учебником, активное чтение, частично-поисковый, исследовательский, эвристическая беседа, метод создания «карт памяти» с элементами коллажирования.
Технологии:
проблемного обучения, информационно-коммуникационные технологии, здоровьесберегающие;
элементы технологии критического мышления и кооперативные методы интерактивного обуч6ения.
Планируемые образовательные результаты
Предметные
Метапредметные
Личностные
- создать организационные и содержательные условия для изучения учащимися микроскопического строения тканей животных;
- содействовать формированию навыка работы с микроскопом и микропрепаратами; умения находить и распознавать виды растительных тканей на рисунках, электронных носителях, микрофотографиях, работать с ЭОР.
- способствовать развитию умения анализировать, сравнивать строение разнообразных тканей животных;
- использовать практические методы познания (наблюдение), рационально используя выделенное время;
- способствовать формированию умения ставить проблему, искать пути ее решения, работать с разнообразными источниками информации для решения проблемы.
- положительное отношение к уроку биологии;
- чувство необходимости учения биологии;
- сформирование познавательных интересов и мотивов, направленных на изучение живой природы;
- осознание необходимости самосовершенствования;
- способность адекватно судить о причинах своего успеха или неуспеха в изучении биологии;
- корректировка своего поведения на уроке биологии в соответствии с моральной нормой.
Основные понятия, изучаемые на уроке
ткани, гистология
Организационная структура урока
№ этапа
Этап урока
УУД
Деятельность
ЭОР
Время,
мин
учителя
учащихся
1
Организационный этап
Регулятивные: волевая саморегуляция, целеполагание как постановка учебной задачи на основе соотнесения того, что уже известно и усвоено учащимся, и того, что еще неизвестно.
Личностные: формирование познавательных интересов и мотивов
-
Приветствие.
-
Анализ готовности учеников класса к уроку (визуально)
-
Наличие ученического методического комплекта
-
Соответствие внешнего вида учащихся требованиям.
-
Психологическая готовность учащихся.
Мотивация учебной деятельности.
Сегодня мы узнаем:
Эта ткань начинает формироваться у эмбриона примерно на 17 день после оплодотворения…
Отростки клеток этой ткани могут достигать размеров в несколько метров.
-
О строении и функциях тканей, которые обеспечивают наше движение, сердцебиение, прохождение пищи через кишечник и др.
-
Об особенностях организации ткани, обеспечивающей регуляцию функций организма и его связь с внешней средой.
Готовятся к уроку, настраивают себя на работу в группах
Лови отгадку
компьютер, мультимедийный проектор
2
2
Этап актуализации субъектного опыта учащихся
Познавательные:
слушать вопросы учителя, выдвигать гипотезы и обосновывать их;
Коммуникативные: планирование учебного сотрудничества с учителем и сверстниками - определение целей, функций участников, способов взаимодействия.
1. Сегодня вам предстоит совершить открытие в области гистологии. Для того, чтобы двигаться дальше в этом направлении, вам необходимо выполнить задание «Найди пару»:
На доске представлены изображения разных типов соединительной и эпителиальных тканей
1 - Костная
А. Связывает кожу с мышцами, удерживая ее в нужном положении, связывает органы
2 -Волокнистая
Б.Входит в состав опорно-двигательной системы, формирует кости
3 - Жировая
В. Ее клетки транспортируют к тканям кислород, а от тканей углекислый газ. Обеспечивает гомеостаз (стабильность внутренней среды организма и защитную функцию)
4 - Кровь
Г. В ее клетках откладываются питательные вещества, защищает органы и организм в целом от механических повреждений и переохлаждения
5 - Хрящевая
Д. Обеспечивает опорную функцию, входит в состав ушных раковин, связок, ОДС.
Давайте вспомним, какие ткани характерны для организма животных? (Эпителий, соединительная, мышечная и нервная ткани).
С какими тканями животных мы уже ознакомились? Какие ткани животных мы еще не знаем?
Что мы на уроке будем изучать? (мышечные и нервную ткани). Записывает тему урока на доске.
Какие задачи мы поставим перед собой на сегодняшнем уроке?
- выяснить, что такое мышечная и нервная ткань?
- рассмотреть ткани в микроскоп;
- - выяснить особенности строения изучаемых тканей;
- выяснить, какие органы животных состоят из этих тканей;
- какое значение имеют эти ткани (их функция).
Запишем план урока на доске:
1. что такое мышечная и нервная ткань?
2. Каковы особенности строения и функции этих тканей.
3 Где локализованы изучаемые ткани
Какими способами мы будем достигать этих целей?
(работая в парах, в группах с микроскопами, самостоятельно в группах с текстом учебника, микрофотографиями)
Слушают учителя.
Выполняют задание, осуществляют взаимопроверку в паре при анализе задания
Выполняя задание, повторяют особенности строения соединительных тканей животных
Воспринимают информацию, сообщаемую учителем.
Отвечают на вопросы учителя. Называют тему урока.
Записывают тему урока в тетради. Ставят перед собой цели.
Отвечают на вопросы учителя.
5
3
Этап изучения новых знаний и способов деятельности
Познавательные:
извлечение необходимой информации, анализ объектов с целью выделения признаков,
обобщения и синтезирования;
Коммуникативные
сотрудничество с учителем и сверстниками.
Личностные:
формировать умение способности к саморазвитию.
1. Образующиеся при делении одинаковые клетки расположены рядом (демонстрация рисунка «Ткани».
Так что же такое ткани? (группы клеток, сходные по строению, происхождению). Из тканей построены различные органы
Почему ткани бывают разные? (потому что выполняют разные функции).
Сегодня вы - исследователи. Работаете в разных лабораториях (лаборатория движения, лаборатория внутренних органов, лаборатория сердца, лаборатория нервного импульса,)
Завершив работу в лабораториях, вам нужно будет ответить на вопрос, почему ваша лаборатория имеет такое название. (В приложении инструктивные карточки).
2. Работа исследователей в лабораториях.
Лабораторная работа №5 «Рассматривание под микроскопом готовых микропрепаратов тканей» по инструктивным карточкам.
-
Заполнение таблицы в рабочей тетради.
Название, схематический рисунок
Особенности строения клеток и межклеточного в-ва
Локализация и особенности функционирования клеток
4. Выступление сотрудников лабораторий в группах (метод «джиг - со»). Обмен информацией. Демонстрация микрофотографий тканейв созданных ИИП с коллажированием.
Воспринимают информацию, сообщаемую учителем, отвечают на вопросы, записывают в тетради
Работают учащиеся в группах - лабораториях.Руководители групп распределяют обязанности между сотрудниками лабораторий. Работают по алгоритму инструктивных карточек.
Заполняют таблицу.
Отчет руководителей лабораторий.
Кооперативный метод интерактивного обучения
15
4
Этап первичной проверки понимания изученного
Познавательные:
анализ объектов с целью выделения признаков,
обобщения и синтезирования.
Личностные: сформированностьздоровьесберегающих мотивов
1. Работа с тренажёром
2. Физкультминутка
Отвечают на вопросы.
Активно участвуют в физкультминутке
5
5
Этап закрепления изученного
Регулятивные: контроль, коррекция, выделение и осознание того, что уже усвоено и что подлежит усвоению, осознание качества и уровня усвоения
Личностные: формирование интеллектуальных умений
Познавательные: поиск и выделение необходимой информации
Выполняют тест.
Вставьте пропущенное слово.
По особенностям строения и функциям узнайте, о какой ткани животного организма идёт речь?
1. Ткань состоит из многоядерных клеток цилиндрической формы , способными к сокращению. Это ____________ ткань.
2. Ткань образована веретеновидными одноядерными клетками, способными к сокращению. Это ____________ ткань.
3. Клетки ткани имеют множество ядер, цилиндрическую форму, переплетены между собой и способны к сокращению. Это ____________ ткань.
4. Ткань образована клетками, способными продуцировать и передавать нервные импульсы, а также вспомогательными клетками. Это ____________ ткань.
5. Высокоспециализированные клетки, имеющие ядро, короткие и длинные отростки, способные генерировать нервный импульс. Это ____________ .
По приведенным признакам обучающиеся узнают ткани, обозначенные цифрами 1-5
Взаимопроверка.
Критерии оценивания: пять правильных ответов - 5, четыре - 4 и т. д.
5
6
Этап обобщения и систематизации
Регулятивные: контроль, коррекция, выделение и осознание того, что уже усвоено, осознание качества и уровня усвоения
Личностные: формирование интеллектуальных умений
Беседа:
- Итак, что такое мышечная «ткань»?
- Какие виды мышечных тканей выделяют у животных?
- Коротко охарактеризуйте каждый тип мышечных тканей.
Что такое нервная ткань?
Что является функциональной единицей нервной ткани?
6
7
Этап информации о домашнем задании
Объясняет сущность домашнего задания. Предлагает дома прочитать п.25, ответить на в.1-4 с.67. Контроль и Итог (пройти тесты) Подготовиться к обобщающему уроку. Творческое задание:
Почему куриное мясо бывает «белое» (например грудинка птицы) и темное, чем они отличаются?
Фиксируют информацию, участвуют в беседе
2
8
Этап подведения итогов учебного занятия
Даёт качественную оценку работы класса и отдельных уч-ся.
Фиксируют выводы
3
9
Рефлексия
Коммуникативные: умения с достаточной полнотой и точностью выражать свои мысли, слушать товарища. Личностные: способность адекватно судить о причинах своего успеха или неуспеха в изучении биологии.
Учитель:
-
Предлагает вспомнить тему и задачи урока
-
Насколько наша деятельность соответствовала теме урока.
-
Продолжить фразу (в тетради)
-
Оценить свою активность на уроке в рамках 10б системы
-
Оценить полезность материала в рамках 10б системы
-
«Хочу заметить что…
Ученики:
- Называют тему и задачи урока
- Отмечают наиболее трудные этапы урока.
- Высказывать оценочные суждения о работе партнера
- Определяют степень совместного продвижения к цели
- Отмечают успешные ответы, интересные вопросы
2
10
Этап «Интересная добавка»
-
«В личный кармашек» секреты кулинарной обработки мяса.
-
«По секрету всему свету» - интересные факты
Мышечные ткани
Мышечные ткани классифицируются на гладкую и исчерченную или поперечнополосатую. Поперечнополосатая подразделяется на скелетную и сердечную. В зависимости от происхождения мышечные ткани делятся на 5 типов:
мезенхимные (гладкая мышечная ткань);
эпидермальные (гладкая мышечная ткань);
нейральные (гладкая мышечная ткань);
целомические (сердечная);
соматические или миотомные (скелетная поперечнополосатая).
ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ, РАЗВИВАЮЩАЯСЯ ИЗ СПЛАНХНОТОМНОЙ МЕЗЕНХИМЫ
локализуется в стенках полых органов (желудка, кровеносных сосудах, дыхательных путях и др.) и неполых органах (в мышце ресничного тела глаза млекопитающих). Клетки гладкой мышечной ткани развиваются из мезенхимоцитов, которые утрачивают отростки. В них развиваются комплекс Гольджи, митохондрии, гранулярная ЭПС и миофиламенты. В это время на гранулярной ЭПС активно синтезируется коллаген V типа, за счет которого вокруг клетки формируется базальная мембрана. При дальнейшей дифференцировке органеллы общего значения атрофируются, снижается синтез молекул коллагена в клетке, но повышается синтез сократительных белков миофиламентов.
СТРОЕНИЕ ГЛАДКОЙ МЫШЕЧНОЙ ТКАНИ. Она состоит из гладких миоцитов, имеющих веретеновидную форму, длиной от 20 до 500 мкм. диаметром 6-8 мкм. Снаружи миоциты покрыты плазмолеммой и базальной мембраной.
Миоциты плотно прилежат друг к другу. Между ними имеются контакты - нексусы. В том месте, где имеются нексусы, в базальной мембране оболочки миоцитов есть отверстия. В этом месте плазмолемма одного миоцита приближается к плазмолемме другого миоцита на расстояние 2-3 нм. Через нексусы происходит обмен ионов, транспорт молекул воды, передача сократительного импульса.
Снаружи миоциты покрыты коллагеном V типа, образующим экзоцитоскелет клетки. Цитоплазма миоцитов окрашивается оксифильно. В ней содержатся слабо развитые органеллы общего значения: гранулярная ЭПС, комплекс Гольджи, гладкая ЭПС, клеточный центр, лизосомы. Эти органеллы располагаются у полюсов ядра. Хорошо развитые органеллы - митохондрии. Ядра имеют палочковидную форму.
В миоцитах хорошо развиты миофиламенты, являющиеся сократительным аппаратом клеток. Среди миофиламентов имеются
тонкие, актиновые, состоящие из белка актина;
толстые миозиновые, состоящие из сократительного белка миозина, которые появляются только после поступления к клетке импульса;
промежуточные филаменты, состоящие из коннектина и небулина.
В миоцитах отсутствует исчерченность потому, что все вышеперечисленные филаменты расположены неупорядоченно.
АКТИНОВЫЕ ФИЛАМЕНТЫ соединяются друг с другом и с плазмолеммой при помощи плотных телец. В тех местах, где они соединяются друг с другом, в тельцах содержится альфа-актинин; в тех местах, где филаменты соединяются с плазмолеммой - в тельцах содержится винкулин. Расположение актиновых филаментов преимущественно продольное, но они могут располагаться под углом по отношению к продольной оси. Миозиновые филаменты тоже располагаются преимущественно продольно. Филаменты располагаются так, что концы актиновых располагаются между концами миозиновых филаментов.
ФУНКЦИЯ ФИЛАМЕНТОВ - сократительная. Процесс сокращения осуществляется следующим образом: после поступления сократительного импульса пиноцитозные пузырьки, содержащие ионы кальция, приближаются к филаментам; ионы кальция запускают сократительный процесс, который заключается в том, что концы актиновых филаментов продвигаются глубже между концами миозиновых филаментов. Сила тяги прилагается к плазмолемме, с которой актиновые филаменты связаны при помощи плотных телец, в результате этого миоцит сокращается.
ФУНКЦИИ МИОЦИТОВ: 1) сократительная (способность к длительному сокращению); 2) секреторная (секретируют коллаген V типа, эластин, протеогликаны, так как имеют гранулярную ЭПС).
РЕГЕНЕРАЦИЯ гладкой мышечной ткани осуществляется 2 путями: 1) митотическое деление миоцитов; 2) преобразование в гладкие миоциты миофибробластов.
СТРОЕНИЕ ГЛАДКОЙ МЫШЕЧНОЙ ТКАНИ КАК ОРГАНА. В стенке полых органов гладкие миоциты образуют пучки. Эти пучки окружены прослойками рыхлой соединительной ткани, которая называется перимизием. Прослойка соединительной ткани вокруг всего пласта мышечной ткани называется эпимизием. В перимизии и эпимизии проходят кровеносные и лимфатические сосуды и нервные волокна.
ИННЕРВАЦИЯ ГЛАДКОЙ МЫШЕЧНОЙ ТКАНИ осуществляется вегетативной нервной системой, поэтому сокращения гладкой мускулатуры не подчиняются воле человека (непроизвольные). К гладкой мышечной ткани подходят чувствительные (афферентные) и двигательные (эфферентные) нервные волокна. Эфферентные нервные волокна заканчиваются двигательными нервными окончаниями в прослойке соединительной ткани. При поступлении импульса из окончаний выделяются медиаторы, которые, диффузно распространяясь, достигают миоцитов, вызывая их сокращение.
ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ ЭПИДЕРМАЛЬНОГО ПРОИСХОЖДЕНИЯ находится в концевых отделах и мелких протоках желез, которые развиваются из кожной эктодермы (слюнные, потовые, молочные и слезные железы). Гладкие миоциты (миоэпителиоциты) располагаются между базальной поверхностью железистых клеток и базальной мембраной, охватывая базальную часть гландулоцитов своими отростками. При сокращении этих отростков сдавливается базальная часть гландулоцитов, благодаря чему из железистых клеток выделяется секрет.
ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ НЕЙРАЛЬНОГО ПРОИСХОЖДЕНИЯ развивается из глазных бокалов, вырастающих из нервной трубки. Эта мышечная ткань образует всего 2 мышцы, расположенные в радужной оболочке глаза: мышцу суживающую зрачок и мышцу расширяющую зрачок. Существует мнение, что мышцы радужки развиваются из нейроглии.
ПОПЕРЕЧНОПОЛОСАТАЯ СКЕЛЕТНАЯ МЫШЕЧНАЯ ТКАНЬ развивается из миотомов мезодермальных сомитов, поэтому называется соматической. Клетки миотомов дифференцируются в двух направлениях: 1) из одних образуются миосателлитоциты; 2) из других образуются миосимпласты.
ОБРАЗОВАНИЕ МИОСИМПЛАСТОВ. Клетки миотомов дифференцируются в миобласты, которые сливаются вместе и образуются мышечные трубочки. В процессе созревания мышечные трубочки превращаются в миосимпласты. При этом ядра смещаются к периферии, а миофибриллы - к центру.
СТРОЕНИЕ МЫШЕЧНОГО ВОЛОКНА. Мышечное волокно (miofibra) состоит из 2 компонентов: 1) миосателлитоцитов и 2) миосимпласта. Мышечное волокно имеет примерно такую же длину, как и сама мышца, диаметр - 20-50 мкм. Волокно снаружи покрыто оболочкой - сарколеммой, состоящей из 2 мембран. Наружная мамбрана называется базальной мембраной, а внутренняя - плазмолеммой. Между этими двумя мембранами располагаются миосателлитоциты.
ЯДРА МЫШЕЧНЫХ ВОЛОКОН располагаются под плазмолеммой, их количество может достигать нескольких десятков тысяч. Имеют вытянутую форму, не обладают способностью к дальнейшему митотическому делению. ЦИТОПЛАЗМА мышечного волокна называется САРКОПЛАЗМОЙ. В саркоплазме содержится большое количество миоглобина, включений гликогена и липидов; имеются органеллы общего значения, одни из которых развиты хорошо, другие - хуже. Такие органеллы как комплекс Гольджи, гранулярная ЭПС, лизосомы развиты слабо и располагаются у полюсов ядер. Хорошо развиты митохондрии и гладкая ЭПС.
В мышечных волокнах хорошо развиты миофибриллы, являющиеся сократительным аппаратом волокна. В миофибриллах имеется исчерченность потому, что миофиламенты в них расположены в строго определенном порядке (в отличии от гладкой мускулатуры). В миофибриллах 2 вида миофиламентов: 1) тонкие актиновые, состоят из белка актина, тропонина и тропомиозина; 2) толстые миозиновые состоят из белка миозина. Актиновые филаменты располагаются продольно, их концы находятся на одинаковом уровне и несколько заходят между концами миозиновых филаментов. Вокруг каждого миозинового филамента расположено 6 концов актиновых филаментов. В мышечном волокне имеется цитоскелет, включающий промежуточные нити (филаменты), телофрагму, мезофрагму, сарколемму. Благодаря цитоскелету одинаковые структуры миофибрилл (актиновые, миозиновые филаменты и др.) располагаются упорядоченно.
Тот участок миофибриллы, в котором находятся только актиновые филаменты, называется диском I (изотропный или светлый диск). Через центр диска I проходит Z-полоска, или телофрагма толщиной около 100 нм и состоящая из альфа-актинина. К телофрагме прикрепляются актиновые нити (зона прикрепления тонких нитей).
Миозиновые филаменты тоже располагаются в строго определенном порядке. Их концы также находятся на одном уровне. Миозиновые филаменты вместе с заходящими между ними концами актиновых филаментов образуют диск А (анизотропный диск, обладающий двулучепреломлением). Диск А также разделяется мезофрагмой, аналогичной телофрагме и состоящей из М-белка (миомизина).
В средней части диска А имеется Н-полоска, ограниченная концами актиновых филаментов, заходящих между концами миозиновых нитей. Поэтому чем ближе концы актиновых филаментов расположены друг к другу, тем уже Н-полоска.
САРКОМЕР - это структурная и функциональная единица миофибрилл, представляющая собой участок, расположенный между двумя телофрагмами. Формула саркомера: 1,5 диска I + диск А + 1,5 диска I. Миофибриллы окружены хорошо развитыми митохондриями и хорошо развитой гладкой ЭПС.
ГЛАДКАЯ ЭПС образует систему L-канальцев, образующих в каждом диске сложные структуры. Эти структуры состоят из L-канальцев расположенных вдоль миофибрилл и соединяющихся с поперечно направленными L-канальцами (латеральными цистернами). ФУНКЦИИ гладкой ЭПС (системы L-канальцев): 1) транспортная; 2) синтез липидов и гликогена; 3) депонирование ионов кальция.
Т-КАНАЛЫ - это впячивания плазмолеммы. На границе дисков из плазмолеммы вглубь волокна происходит впячивание в виде трубочки, располагающейся между двумя латеральными цистернами.
ТРИАДА включает: 1) Т-канал и 2) 2 латеральные цистерны гладкой ЭПС. ФУНКЦИЯ ТРИАД заключается в том, что в расслабленном состоянии миофибрилл в латеральных цистернах накапливаются ионы кальция; в тот момент, когда по плазмолемме движется импульс (потенциал действия), он переходит на Т-каналы. При движении импульса по Т-каналу из латеральных цистерн выходят ионы кальция. Без ионов кальция невозможно сокращение миофибрилл, потому что в актиновых филаментах центры взаимодействия с миозиновыми нитями заблокированы тропомиозином. Ионы кальция осуществляют разблокирование этих центров, после чего начинается взаимодействие актиновых нитей с миозиновыми и начинается сокращение.
МЕХАНИЗМ СОКРАЩЕНИЯ МИОФИБРИЛЛ. При взаимодействии актиновых филаментов с миозиновыми происходит разблокирование ионами Са центров сцепления актиновых филаментов с головками молекул миозина, после чего эти выросты присоединяются к центрам сцепления на актиновых нитях и как веслом осуществляют движение актиновых филаментов между концами миозиновых. В это время телофрагма приближается к концам миозиновых филаментов, поскольку концы актиновых филаментов тоже приближаются к мезофрагме и друг к другу, постольку происходит сужение Н-полоски. Таким образом, во время сокращения миофибрилл происходит сужение диска I и Н-полоски. После прекращения потенциала действия ионы кальция возвращаются в L-канальцы гладкой ЭПС, тропомиозин снова блокирует в актиновых филаментах центры взаимодействия с миозиновыми нитями. Это приводит к прекращению сокращения миофибрилл, происходит их расслабление, т.е. актиновые нити возвращаются в исходное положение, восстанавливается ширина диска I и Н-полоски.
МИОСАТЕЛЛИТОЦИТЫ мышечного волокна располагаются между базальной мембраной и плазмолеммой сарколеммы. Эти клетки имеют овальную форму, их овальное ядро окружено тонким слоем бедной органеллами и слабо окрашиваемой цитоплазмы. ФУНКЦИЯ миосателлитоцитов - это камбиальные клетки, участвующие в регенерации мышечных волокон при их повреждении.
СТРОЕНИЕ МЫШЦЫ КАК ОРГАНА. Каждая мышца тела человека представляет собой своеобразный орган, имеющий свою структуру. Каждая мышца состоит из мышечных волокон. Каждое волокно окружено тонкой прослойкой рыхлой соединительной ткани - эндомизием. В эндомизии проходят кровеносные и лимфатические сосуды и нервные волокна. Мышечное волокно вместе с сосудами и нервными волокнами называется "мион". Несколько мышечных волокон образуют пучок, окруженный слоем рыхлой соединительной ткани, называемой перимизием. Вся мышца окружена прослойкой соединительной ткани, называемой эпимизием.
СВЯЗЬ МЫШЕЧНЫХ ВОЛОКОН С КОЛЛАГЕНОВЫМИ ВОЛОКНАМ СУХОЖИЛИЙ.
На концах мышечных волокон имеются впячивания сарколеммы. В эти впячивания входят коллагеновые и ретикулярные волокна сухожилий. Ретикулярные волокна прободают базальную мембрану и при помощи молекулярных сцеплений соединяются с плазмолеммой. Затем эти волокна возвращаются в просвет впячивания и оплетают коллагеновые волокна сухожилия, как бы привязывая их к мышечному волокну. Коллагеновые волокна образуют сухожилия, которые прикрепляются к костному скелету.
ТИПЫ МЫШЕЧНЫХ ВОЛОКОН. Имеется 2 основных типа мышечных волокон:
I тип (красные волокна) и II тип (белые волокна). Они различаются главным образом по быстроте сокращения, содержанию миоглобина, гликогена и активности ферментов.
1-й ТИП (красные волокна) характеризуются большим содержанием миоглобина (поэтому они красные), высокой активностью сукцинатдегидрогеназы, АТФ-азой медленного типа, не столь богатым содержанием гликогена, длительностью сокращения и малой утомляемостью.
2-й ТИП (белые волокна) отличаются малым содержанием миоглобина, низкой активностью сукцинатдегидрогеназы, АТФ-азой быстрого типа, богатым содержанием гликогена, быстрым сокращением и большой утомляемостью.
Медленный (красный) и быстрый (белый) тип мышечных волокон иннервируются разными типами моторных нейронов: медленным и быстрым. Кроме 1-го и 2-го типов мышечных волокон имеются промежуточные, обладающие свойствами тех и других.
В каждой мышце имеются все типы мышечных волокон. Их количество может меняться и зависит от физической нагрузки.
РЕГЕНЕРАЦИЯ ПОПЕРЕЧНОПОЛОСАТОЙ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ. При повреждении (разрыве) мышечных волокон их концы на месте повреждения подвергаются некрозу. После разрыва к обрывкам волокон поступают макрофаги, которые фагоцитируют некротизированные участки, очищая их от мертвой ткани. После этого процесс регенерации осуществляется 2 путями: 1) за счет повышения реактивности в мышечных волокнах и образования мышечных почек в местах разрыва; 2) за счет миосателлитоцитов.
1-й ПУТЬ характеризуется тем, что на концах разорванных волокон гипертрофируется гранулярная ЭПС, на поверхности которой синтезируются белки миофибрилл, мембранных структур внутри волокна и сарколеммы. В результате этого концы мышечных волокон утолщаются и преобразуются в мышечные почки. Эти почки по мере их увеличения приближаются друг к другу от одного оборванного конца к другому, наконец почки соединяются и срастаются. Между тем за счет клеток эндомизия происходит новообразование соединительной ткани между растущими навстречу друг к другу мышечными почками. Поэтому к моменту соединения мышечных почек формируется соединительнотканная прослойка, которая войдет в состав мышечного волокна. Следовательно, формируется соединительнотканный рубец.
2-й ПУТЬ регенерации заключается в том, что миосателлитоциты покидают места своего обитания и подвергаются дифференцировке, в результате которой превращаются в миобласты. Часть миобластов присоединяется к мышечным почкам, часть соединяется в мышечные трубочки, которые дифференцируются в новые мышечные волокна.
Таким образом, при репаративной регенерации мышц восстанавливаются старые мышечные волокна и образуются новые.
ИННЕРВАЦИЯ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ осуществляется двигательными и чувствительными нервными волокнами, заканчивающимися нервными окончаниями. ДВИГАТЕЛЬНЫЕ (моторные) нервные окончания являются концевыми приборами аксонов моторных нервных клеток передних рогов спинного мозга. Конец аксона, подходя к мышечному волокну делится на несколько веточек (терминалей). Терминали прободают базальную мембрану сарколеммы и далее погружаются вглубь мышечного волокна, увлекая за собой плазмолемму. В результате этого образуется нервномышечное окончание (моторная бляшка).
СТРОЕНИЕ НЕРВНОМЫШЕЧНОГО окончания. В нервномышечном окончании имеется две части (полюса): нервная и мышечная. Между нервной и мышечной частями имеется синаптическая щель. В нервной части (терминалях аксона моторного нейрона) имеются митохондрии и синаптические пузырьки, заполненные медиатором-ацетилхолином. В мышечной части нервномышечного окончания есть митохондрии, скопление ядер, отсутствуют миофибриллы. Синаптическая щель шириной 50 нм ограничена пресинаптической мембраной (плазмолеммой аксона) и постсинаптической мембраной (плазмолеммой мышечного волокна). Постсинаптическая мембрана образует складки (вторичные синаптические щели), на ней имеются рецепторы к ацетилхолину и фермент - ацетилхолинэстераза..
ФУНКЦИЯ нервно-мышечных окончаний. Импульс движется по плазмолемме аксона (пресинаптической мембране). В это время синаптические пузырьки с ацетилхолином подходят к плазмолемме, из пузырьков ацетилхолин изливается в синаптическую щель и захватывается рецепторами постсинаптической мембраны. Это повышает проницаемость этой мембраны (плазмолеммы мышечного волокна), в результате этого ионы натрия с наружной поверхности плазмолеммы переходят на внутреннюю, а ионы калия переходят на наружную поверхность - это и есть волна деполяризации или нервный импульс (потенциал действия). После возникновения потенциала действия ацетилхолинэстераза постсинаптической мембраны разрушает ацетилхолин и прекращается переход импульса через синаптическую щель.
ЧУВСТВИТЕЛЬНЫМИ НЕРВНЫМИ ОКОНЧАНИЯМИ (нервно-мышечными веретенами - fusi neuro-muscularis) заканчиваются дендриты чувствительных нейронов спинномозговых узлов. Нервно-мышечные веретена покрыты соединительнотканной капсулой, внутри которой имеются 2 типа интрафузальных (внутриверетенных) мышечных волокон: 1) с ядерной сумкой (в центре волокна утолщение, в котором имеется скопление ядер), они более длинные и более толстые; 2) с ядерной цепочкой (ядра ввиде цепочки располагаются по центру волокна), они тоньше и короче.
В окончания проникают толстые нервные волокна, которые кольцеобразно оплетают оба вида интрафузальных мышечных волокон и тонкие нервные волокна, заканчивающиеся гроздъевидными окончаниями на мышечных волокнах с ядерной цепочкой. На концах интрафузальных волокон имеются миофибриллы и к ним подходят двигательные нервные окончания. Сокращения интрафузальных волокон не обладают большой силой и не суммируются с остальными (экстрафузальными) волокнами мышцы.
ФУНКЦИЯ нервно-мышечных веретен заключается в восприятии скорости и силы растяжения мышцы. Если сила растяжения такова, что угрожает разрывом мышцы, то на сокращающиеся мышцы-антогонисты от этих окончаний рефлекторно поступают тормозные импульсы.
СЕРДЕЧНАЯ МЫШЕЧНАЯ ТКАНЬ развивается из переднего отдела висцеральных листков спланхнотома. Из этих листков выделяются 2 миоэпикардиальных пластинки: правая и левая. Клетки миоэпикардиальных пластинок дифференцируются в двух направлениях: из одних развивается мезотелий, покрывающий эпикард, из других - кардиомиоциты пяти разновидностей;
сократительные
пейсмекерные
проводящие
промежуточные
секреторные, или эндокринные
.
СТРОЕНИЕ КАРДИОМИОЦИТОВ. Кардиомиоциты имеют цилиндрическую форму, длиной 50-120 мкм, диаметром 10-20 мкм. Кардиомиоциты соединяются концами друг с другом и образуют функциональные сердечные мышечные волокна. Места соединения кардиомиоцитов называются вставочными дисками (discus intercalatus). В дисках имеются интердигитации, десмосомы, места прикрепления актиновых филаментов и нексусы. Через нексусы происходит обмен веществ между кардиомиоцитами.
Снаружи кардиомиоциты покрыты сарколеммой, состоящей из наружной (базальной) мембраны и плазмолеммы. От боковых поверхностей кардиомиоцитов отходят отростки, вплетающиеся в боковые поверхности кардиомиоцитов соседнего волокна. Это мышечные анастомозы.
ЯДРА кардиомиоцитов (одно-два), овальной формы, обычно полиплоидные, располагаются в центре клетки. МИОФИБРИЛЛЫ локализованы по периферии. ОРГАНЕЛЛЫ - одни развиты слабо (гранулярная ЭПС, комплекс Гольджи, лизосомы), другие - хорошо (митохондрии, гладкая ЭПС, миофибриллы). В оксифильной ЦИТОПЛАЗМЕ имеются включения миоглобина, гликогена и липидов.
СТРОЕНИЕ МИОФИБРИЛЛ такое же как и в скелетной мышечной ткани. Актиновые филаменты формируют светлый диск ( I ), разделенный телофрагмой, за счет миозиновых филаментов и концов актиновых образуется диск А (анизотропный), разделенный мезофрагмой. В средней части диска А имеется Н-полоска, ограниченная концами актиновых филаментов.
Волокна сердечной мышцы отличаются от волокон скелетной мускулатуры тем, что состоят они из отдельных клеток - кардиомиоцитов, наличием мышечных анастомозов, центральным расположением ядер (в волокне скелетной мышцы - под сарколеммой), увеличенной толщиной диаметра Т-каналов, так как в их состав входит и плазмолемма и базальная мембрана (в волокнах скелетной мышцы - только плазмолемма).
ПРОЦЕСС СОКРАЩЕНИЯ в волокнах сердечной мышцы осуществляется по такому же принципу, как и волокнах скелетной мышечной ткани.
ПРОВОДЯЩИЕ КАРДИОМИОЦИТЫ характеризуются более толстым диаметром (до 50 мкм), более светлой цитоплазмой, центральным или эксцентричным расположением ядер, малым содержанием миофибрилл, более простым устройством вставочных дисков. В дисках меньше десмосом, интердигитаций, нексусов и мест прикрепления актиновых филаментов.
В проводящих кардиомиоцитах отсутствуют Т-каналы. Проводящие кардиомиоциты могут соединяться друг с другом не только своими концами, но и боковыми поверхностями. ФУНКЦИЯ проводящих кардиомиоцитов заключается в выработке и передаче сократительного импульса на сократительные кардиомиоциты.
ЭНДОКРИННЫЕ КАРДИОМИОЦИТЫ располагаются только в предсердиях, имеют более отростчатую форму, слабо развитые миофибриллы, вставочные диски, Т-каналы. В них хорошо развиты гранулярная ЭПС, комплекс Гольджи и митохондрии, в их цитоплазме имеются гранулы секрета.
ФУНКЦИЯ эндокринных кардиомиоцитов - секреция предсердного натрийуретического фактора (ПНФ), который регулирует сократимость сердечной мышцы, объем циркулирующей жидкости, артериальное давление, диурез.
РЕГЕНЕРАЦИЯ сердечной мышечной ткани только физиологическая, внутриклеточная. При повреждении волокон сердечной мышцы, они не восстанавливаются, а замещаются соединительной тканью (гистотипическая регенерация).
Нервная ткань
Химические основы жизнедеятельности нервной ткани имеют, с одной стороны, общие черты, присущие клеткам любой ткани, с другой специфические особенности, определяемые характером функций, выполняемых нервной системой в целостном организме. Эти особенности проявляются как в химическом составе, так и в метаболизме нервной ткани.
Нервная ткань состоит из трех клеточных элементов: нейронов (нервных клеток); нейроглии - системы клеток, непосредственно окружающих нервные клетки в головном и спинном мозге; мезенхимных элементов, включающих микроглию (клетки Гортега).
Основная масса головного мозга - это первые два типа клеточных элементов. Нейроны сосредоточены в сером веществе (60-65% от вещества головного мозга), тогда как белое вещество ЦНС и периферические нервы состоят главным образом из элементов нейроглии и их производного - миелина.
СТРУКТУРА НЕЙРОНА
Нейрон состоит из тела клетки, многочисленных ветвящихся коротких отростков - дендритов и одного длинного отростка - аксона, длина которого может достигать нескольких десятков сантиметров (рис. 134).
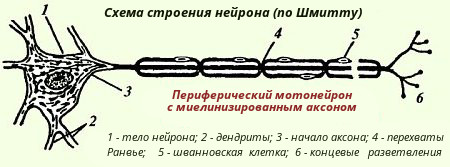
Объем цитоплазмы, содержащейся в отростках нервной клетки, может в несколько раз превышать ее количество в теле клетки. Тело нейрона окружено плазматической мембраной - плазмалеммой (рис. 135). В тесной связи с плазмалеммой1 в теле нейрона и проксимальных отрезках дендритов находится так называемая подповерхностная мембранная структура. Это - цистерны, которые расположены параллельно поверхности плазмалеммы и отделены от нее очень узкой светлой зоной. (1При возбуждении нейрона проницаемость плазматической мембраны из меняется.)
Предполагают, что цистерны играют важную роль в метаболизме нейрона. Основной ультраструктурой цитоплазмы нейрона является эндоплазматический ретикулум (или эргастоплазматическая сеть). Эндоплазматический ретикулум - система ограниченных мембраной пузырьков, трубочек и уплощенных мешочков, или цистерн. Мембраны эндоплазматического ретикулума связаны определенным образом с плазмалеммой и оболочкой ядра нейрона.
Гранулы, локализованные на мембранах эндоплазматического ретикулума, а также свободно расположенные в цитоплазме, являются рибосомами.
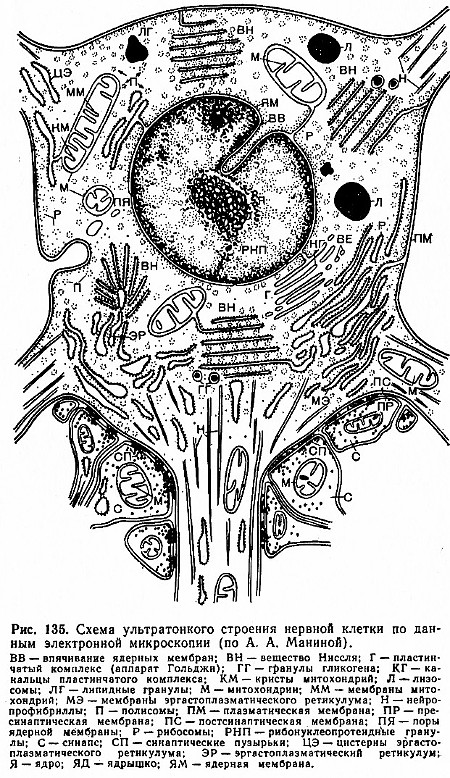
Характерной структурной единицей нервной клетки являются тельца Ниссля, состоящие из рибонуклеиновых кислот и белков. В цитоплазме также выявляется сеть тонких нитей - нейрофибрилл, которые в совокупности образуют густую сеть. Нейрофибриллы - это структурное выражение правильной линейной ориентации белковых молекул.
Важный компонент цитоплазмы нейрона - пластинчатый комплекс (аппарат Гольджи), где сосредоточены главным образом липидные компоненты клетки. Одной из особенностей митохондрий, изолированных из нервных клеток, является то, что они содержат меньше ферментов, участвующих в процессах окисления жирных кислот и аминокислот, чем митохондрии из других тканей.
В ЦНС лизосомы обнаруживаются постоянно и выполняют те же функции, что и лизосомы других органов и тканей.
Размер ядра нейрона колеблется от 3 до 18 мкм, достигая в крупных нейронах 1/4 величины их тела.
СТРОЕНИЕ МИЕЛИНА
Нервные волокна, образующиеся из аксонов нервных клеток, по своему строению могут быть подразделены на два типа: мякотные (миелиновые) и безмякотные (бедные миелином).
Проводниковая система соматической нервной системы, а также ЦНС относится к первому типу, функционально более совершенному, обладающему способностью с высокой скоростью передавать нервные импульсы.
Миелиновое вещество - понятие чисто морфологическое. По сути миелин - система, образованная многократно наслаивающимися мембранами клеток нейроглии2 вокруг нервных отростков (в периферических нервных стволах нейроглия представлена леммоцитами, или шванновскими клетками, а в белом веществе ЦНС - астроцитами макроглии). (2Тонкая структура нейроглии рассматривается в специальных руководствах, посвященных гистологии и морфологии нервной системы.)
По химическому составу миелиновое вещество является сложным белково-липидным комплексом. На долю липидов приходится до 80% плотного остатка; 90% всех липидов миелина представлено холестерином, фосфолипидами и цереброзидами.
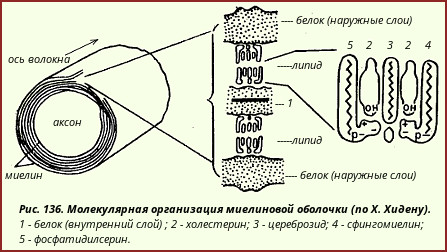
Есть основания полагать, что в липоидных слоях миелиновых оболочек молекулы различных липидов имеют строго определенное расположение (рис. 136).
ХИМИЧЕСКИЙ СОСТАВ ГОЛОВНОГО МОЗГА
Серое вещество головного мозга представлено в основном телами нейронов, а белое вещество - аксонами. В связи с этим указанные отделы мозга значительно отличаются по своему химическому составу. Эти отличия носят прежде всего количественный характер. Содержание воды в сером веществе головного мозга заметно больше, чем в белом веществе (табл. 54).
Таблица 54. Химический состав серого и белого вещества головного мозга человека (в процентах от массы сырой ткани)
Составные части
Серое вещество
Белое вещество
Вода
84
70
Сухой остаток
16
30
Белки
8
9
Липиды
5
17
Минеральные вещества
1
2
В сером веществе белки составляют половину плотных веществ, а в белом веществе - одну треть3. На долю липидов в белом веществе приходится более половины сухого остатка, в сером веществе - лишь около 30%. (3При расчете на сырую массу ткани белкн распределяются примерно поровну между серым (8%) и белым (9%) веществом головного мозга.)
Белки головного мозга
На долю белков приходится примерно 40% сухой массы головного мозга. Мозговая ткань является трудным объектом для изучения белкового состава вследствие большого содержания липидов и наличия белково-липидных комплексов.
Впервые А. Я. Данилевский разделил белки мозговой ткани на растворимые в воде и солевых растворах белки и нерастворимые белки. Обширные исследования в этой области были проведены также А. В. Палладиным и сотр., которые разделили белки нервной ткани на четыре фракции: извлекаемые водой, 4,5% раствором КС1, 0,1% раствором NaOH и нерастворимый остаток. Установлено, что серое вещество богаче белками, растворимыми в воде, чем белое вещество: соответственно 30% и 19%. Белое вещество, напротив, содержит гораздо больше (22%) нерастворимого белкового остатка, чем серое вещества (5%).
В дальнейшем было выделено 5-10 фракций растворимых белков мозга, отличающихся по своей электрофоретической подвижности.
В настоящее время, сочетая методы экстракции буферными растворами, хроматографии на колонках с ДЭАЭ-целлюлозой и диск-электрофореза в полиакриламидном геле, удалось из ткани мозга выделить около 100 различных растворимых белковых фракций.
В нервной ткани содержатся как простое, так и сложные белки. Простые белки - альбумины (нейроальбумины), глобулины (нейроглобулины), катионные белки (гистоны и др.) и опорные белки (нейросклеропротеиды).
Поскольку альбумины и глобулины головного мозга по своим физико-химическим свойствам несколько отличаются от аналогичных белков сыворотки крови, они, как правило, называются нейроальбуминами и нейроглобулинами. Количество нейроглобулинов в головном мозге относительно невелико - в среднем 5% по отношению ко всем растворимым белкам. Нейроальбумины являются основным белковым компонентом фосфопротеидов нервной ткани, на их долю приходится основная масса растворимых белков (89-90%). В свободном состоянии нейроальбумины встречаются редко. Большая часть нейроглобулинов входит в состав сложных белков. В частности, они легко соединяются с липидами, нуклеиновыми кислотами, углеводами и другими небелковыми компонентами.
Белки, которые в процессе электрофоретического разделения при pH 10,5-12,0 движутся к катоду, получили название катионных. Главнейшими представителями этой группы белков в нервной ткани являются гистоны, которые делятся на пять основных фракций в зависимости от содержания в их полипептидных цепях остатков лизина, аргинина и глицина.
Нейросклеропротеиды можно охарактеризовать как структурно-опорные белки. Основные представители этих белков - нейроколлагены, нейроэластины, нейростромины и др. Они составляют примерно 8-10% от всех простых белков нервной ткани и локализованы в основном в белом веществе головного мозга и в периферической нервной системе.
Сложные белки нервной ткани представлены нуклеопротеидами, липопротеидами, протеолипидами, фосфопротеидами, гликопротеидами и т. д. В мозговой ткани содержатся в значительном количестве еще более сложные надмолекулярные образования, такие, как липонуклеопротеиды, липогликопротеиды, возможно, и липогликонуклеопротеидные комплексы.
Нуклеопротеиды - белки, которые принадлежат либо к дезоксирибонуклеопротеидам (ДНП), либо к рибонуклеопротеидам (РНП). Часть этих белков из мозговой ткани извлекается водой, другая часть - солевыми средами, а третья - 0,1 н. раствором щелочи.
Липопротеиды имеют в своем составе липидные группировки. Они составляют значительную часть водорастворимых белков мозговой ткани. Их липидный компонент состоит в основном из фосфолипидов и холестерина.
Протеолипиды - единственные сложные белки, которые извлекаются органическими растворителями, например, смесью хлороформа и метанола. В отличие от липопротеидов в них липидный компонент преобладает над белковым. Наибольшее количество протеолипидов сосредоточено в миелине, в небольших количествах они входят в состав синаптических мембран и синаптических пузырьков.
Фосфопротеиды - сложные белки, простетической группой которых чаще является фосфатная группа, соединенная по типу сложноэфирной связи с остатком серина. Содержание фосфопротеидов в головном мозге более высокое, чем в других органах и тканях, - около 2% по отношению ко всем сложным белкам мозга. Фосфопротеиды обнаружены в мембранах различных морфологических структур нервной ткани.
Гликопротеиды представляют собой чрезвычайно гетерогенную группу белков. По количеству белка и углеводов, входящих в состав гликопротеидов, их можно разделить на две основные группы. Первая группа - это гликопротеиды, в которых ох 5 до 40% углеводов и их производных; белковая часть состоит преимущественно из альбуминов и глобулинов. В гликопротеидах, составляющих вторую группу, содержится 40-85%. углеводов, часто обнаруживается липидный компонент; по своему составу они могут быть отнесены к гликолипопротеидам.
В последние годы в нервной ткани обнаружен ряд специфических белков. К таким белкам, в частности, относятся белок S-100 и белок 14-3-2. Белок S-100, или белок Мура, называют также кислым белком, так как он содержит большое количество остатков глутаминовой и аспарагиновой кислот. Этот белок сосредоточен в основном в нейроглии (85-90%), в нейронах его не более 10-15% от общего количества в головном мозге. Установлено, что концентрация белка S-100 возрастает при обучении (тренировках) животных. Однако пока нет оснований считать, что белок S-100 непосредственно участвует в формировании и хранении памяти. Не исключено, что его участие в этих процессах опосредованное.
Белок 14-3-2 также относится к кислым белкам. В отличие от белка S-100 он локализован в основном в нейронах; в нейроглиальных клетках его содержание невелико. Пока неясна роль белка 14-3-2 в выполнении специфических функций нервной ткани.
Ферменты. В мозговой ткани содержится большое количество ферментов, катализирующих обмен углеводов, липидов и белков. Однако до сих пор в кристаллическом виде из ЦНС млекопитающих выделены лишь два фермента - ацетилхолинэстераза и креатинкиназа.
Значительное количество ферментов в мозговой ткани находится в нескольких молекулярных формах (изоферменты): лактатдегидрогеназа, альдолаза, креатинкиназа, гексокиназа, малатдегидрогеназа, глутаматдегидрогеназа, холинэстераза, кислая фосфатаза, моноаминоксидаза и другие.
Таблица 55. Липидный состав нервной ткани (по Norton, Poduslo, Suzuki, 1966)
Серое вещество
Белое вещество
Миелин
Общее содержание липидов, % от сухой массы
32,7
54,9
70
В процентах к общим липидам
Холестерин
22,0
27,5
27,7
Цереброзиды
5,4
19,8
22,7
Ганглиозиды
1,7
5,4
3,8
Фосфатидилэтаноламины
22,7
14,9
15,6
Фосфатидилхолины
26,7
12,8
11,2
Фосфатидилсерины
8,7
7,9
4,8
Фосфатидилинозиты
2,7
0,9
0,6
Плазмалогены
8,8
11,2
12,3
Сфингомиелины
6,9
7,7
7,9
Липиды
Среди химических компонентов головного мозга особое место занимают липиды, высокое содержание и специфическая природа которых придают мозговой ткани характерные особенности. В группу липидов головного мозга входят фосфолипиды, холестерин, сфингомиелины, цереброзиды, ганглиозиды и очень небольшое количество нейтрального жира (табл. 55). Кроме того, многие липиды нервной ткани находятся в тесной взаимосвязи с белками, образуя, в частности, сложные системы типа протеолипидов.
В сером веществе головного мозга фосфолипиды составляют более 60% от всех липидов, а в белом веществе - около 40%. Напротив, в белом веществе содержание холестерина, сфингомиелинов и особенно цереброзидов больше, чем в сером веществе.
Углеводы
В мозговой ткани имеются гликоген и глюкоза. Однако по сравнению с другими тканями ткань мозга бедна углеводами. Общее содержание глюкозы в головном мозге разных животных составляет в среднем 1-4 мкмоля на 1 г ткани, а гликогена - 2,5-4,5 мкмоля на 1 г ткани (в расчете на глюкозу). Интересно отметить, что общее содержание гликогена в мозге эмбрионов и новорожденных животных значительно выше, чем в мозге взрослых. Например, у новорожденных мышей в отличие от взрослых особей уровень гликогена в 3 раза выше. По мере роста и дифференцировки мозга концентрация гликогена быстро снижается и остается относительно постоянной у взрослого животного.
В мозговой ткани имеются также промежуточные продукты обмена углеводов: гексозо- и триозофосфаты, молочная, пировиноградная и другие кислоты. В табл. 56 приведены данные о содержании некоторых промежуточных компонентов обмена углеводов в головном мозге крыс.
Таблица 56. Средние данные о содержании некоторых метаболитов обмена углеводов в головном мозге крыс, (по Bergmeyer, 1970)
Метаболит
Содержание, мкмоль на 1 г сырой массы ткани
Глюкозо-6-фосфат
0,039-0,049
Фруктозо-6-фосфат
0,017-0,023
Фруктозо-1,6-дифосфат
0,010-0,017
Фосфодиоксиацетон
0,024
3-Фосфоглицериновый альдегид
0,021-0,046
3-Фосфоглицерат
0,085-0,100
2-Фосфоглицерат
0,010-0,016
Фосфоенолпируват
0,035-0,097
Пируват
0,120-0,190
Лактат
1,26-1,70
Адениновые нуклеотиды и креатинфосфат
Из свободных нуклеотидов в мозговой ткани на долю адениновых нуклеотидов приходится около 84%. Большую часть оставшихся нуклеотидов составляют производные гуанина. В целом количество макроэргических соединений в нервной ткани невелико. Так, в головном мозге крыс содержание нуклеотидов и креатинфосфата в среднем составляет (в мкмоль на 1 г сырой массы): АТФ-2,30-2,90; АДФ-0,30-0,50; АМФ-0,03-0,05; ГТФ-0,20-0,30; ГДФ-0,15-0,20; УТФ-0,17-0,25; креатинфосфат - 3,50-4,75. Распределение основных макроэргическил соединений примерно одинаково во всех отделах мозга.
Содержание циклических нуклеотидов (цАМФ и цГМФ) в головном мозге значительно выше, чем во многих других тканях. Уровень цАМФ в мозге составляет в среднем 1-2, а цГМФ-до 0,2 нмоль на 1 г ткани. Для мозга характерна также и высокая активность ферментов метаболизма циклических нуклеотидов. Большинство исследователей считают, что циклические нуклеотиды участвуют в синаптической передаче.
Минеральные вещества
Na, К, Си, Fe, Са, Mg и Мn распределены в головном мозге относительно равномерно между серым и белым веществом. Содержание же фосфора в белом веществе выше, чем в сером.
В табл. 57 представлены средние данные о содержании основных минеральных компонентов в головном мозге и биожидкостях организма.
Таблица 57. Содержание основных минеральных компонентов в ткани головного мозга и плазме крови человека
Компонент
Мозговая ткань, ммоль/кг
Плазма крови, ммоль/л
Na+
57
141
К+
96
5
Са2+
1
2,5
Cl-
37
101
НСО-3
12
28
Как видно из данных, приведенных в табл. 59, концентрация ионов калия, натрия, а также хлора в мозге резко отличается от концентрации их в жидкостях тела.
Количественное соотношение неорганических анионов и катионов в мозговой ткани свидетельствует о дефиците анионов. Расчет показывает, что для покрытия дефицита анионов потребовалось бы в 2 раза больше белков, чем их имеется в мозговой ткани. Принято считать, что остающийся дефицит анионов покрывается за счет липидов. Вполне возможно, что участие липидов в ионном балансе - одна из функций липидов в деятельности головного мозга.
ОСОБЕННОСТИ МЕТАБОЛИЗМА НЕРВНОЙ ТКАНИ
-
Дыхание [показать]
-
Обмен глюкозы и гликогена в головном мозге [показать]
-
Обмен лабильных фосфатов (макроэргов) в ткани мозга [показать]
-
Обмен белков и аминокислот [показать]
-
Обмен липидов [показать]
ХИМИЧЕСКИЕ ОСНОВЫ ВОЗНИКНОВЕНИЯ И ПРОВЕДЕНИЯ НЕРВНЫХ ИМПУЛЬСОВ
Каковы же химические основы возникновения и поддержания биоэлектрических потенциалов (потенциалов покоя и действия)? Большинство исследователей придерживаются мнения, что явления электрической поляризации клетки обусловлены неравномерным распределением ионов калия и натрия по обе стороны клеточной мембраны. Мембрана обладает избирательной проницаемостью: большей для ионов калия и значительно меньшей для ионов натрия. Кроме того, в нервных- клетках существует механизм, который поддерживает внутриклеточное содержание натрия на низком уровне вопреки градиенту концентрации. Этот механизм получил название "натриевого насоса".
При определенных условиях резко повышается проницаемость мембраны для ионов натрия.
В состоянии покоя внутренняя сторона клеточной мембраны заряжена электроотрицательно по отношению к наружной поверхности. Объясняется это тем, что количество ионов натрия, выкачиваемых из клетки с помощью "натриевого насоса", не вполне точно уравновешивается поступлением в клетку ионов калия. В связи с этим часть катионов натрия удерживается внутренним слоем противоионов (анионов) на наружной поверхности клеточной мембраны.
При возбуждении, вызванном тем или иным агентом, селективно изменяется проницаемость мембраны нервной клетки (аксона): увеличивается избирательно для ионов натрия (примерно в 500 раз) и остается без изменения для ионов калия. В результате ионы натрия устремляются внутрь клетки. Компенсирующий поток ионов калия, направляющийся наружу из клетки, несколько запаздывает. Это приводит к возникновению отрицательного заряда на наружной поверхности клеточной мембраны. Внутренняя поверхность мембраны приобретает положительный заpяд; происходит перезарядка клеточной мембраны (в частности, мембраны аксона, т. е. нервного волокна) и возникает потенциал действия, или спайк. Продолжительность спайка не превышает 1 мс. Он имеет восходящую фазу, пик и нисходящую фазу. Нисходящая фаза (падение потенциала) связана с нарастающим преобладанием выхода ионов калия над поступлением ионов натрия - мембранный потенциал возвращается к норме. После проведения импульса в клетке восстанавливается состояние покоя. В этот период ионы натрия, вошедшие в нейрон при возбуждении, заменяются на ионы калия. Этот переход происходит против градиента концентрации, так как ионов натрия во внешней среде, окружающей нейроны, намного больше, чем в клетке после момента ее возбуждения. Переход ионов натрия против градиента концентрации, как уже отмечалось, осуществляется с помощью натриевого насоса, для работы которого необходима энергия АТФ. В конце концов все это приводит к восстановлению исходной концентрации катионов калия и натрия внутри клетки (аксона), и нерв готов для получения следующего импульса возбуждения.
Другим не менее важным процессом для нервной ткани является передача нервного импульса от одной нервной клетки к другой или воздействие на клетки эффекторного органа.
Роль медиаторов в передаче нервных импульсов
В мозге есть миллиарды нейронов, общающихся друг с другом посредством медиаторов.
Химическое вещество можно отнести к числу медиаторов лишь в том случае, если оно удовлетворяет ряду критериев. В нервных волокнах должны содержаться ферменты, необходимые для синтеза этого вещества. При раздражении нервов это вещество должно выделяться, реагировать со специфическим рецептором на постсинаптической клетке и вызывать биологическую реакцию. Должны существовать механизмы, быстро прекращающие действие этого химического вещества.
Всем этим критериям удовлетворяют два вещества - ацетилхолин и норадреналин. Содержащие их нервы называют соответственно холинергическими и адренергическими. В соответствии с этим все эфферентные системы делят на холинореактивные и адренореактивные.
Ряд других химических веществ удовлетворяет многим, но не всем перечисленным критериям. К таким медиаторам относятся дофамин, адреналин, серотонин, октопамин, гистамин, γ-аминомасляная кислота и др.
Обширная группа холинореактивных систем весьма неоднородна как в структурном, так и в функциональном отношении. Объединяют эти системы медиатор - ацетилхолин - и общая схема строения синапса.
Ацетилхолин представляет собой сложный эфир уксусной кислоты и холина. Он синтезируется в нервной клетке из холина и активной формы ацетата - ацетилкоэнзима А при помощи специального фермента холинацетилтрансферазы (холинацетилазы):

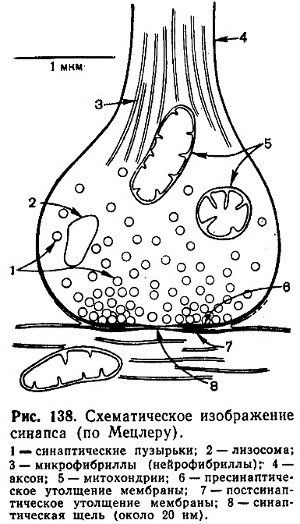
Синапс можно представить себе как узкое пространство - щель, ограниченное с одной стороны пресинаптической, с другой - постсинаптической мембраной (рис. 138). Пресинаптическая мембрана состоит из внутреннего слоя, принадлежащего цитоплазме нервного окончания, и наружного слоя, образованного нейроглией. Мембрана в некоторых местах утолщена и уплотнена, в других - истончена и имеет отверстия, с помощью которых цитоплазма аксона может сообщаться с синаптическим пространством. Постсинаптическая мембрана менее плотная, не имеет отверстий. Подобным образом построены и нервно-мышечные синапсы, но они имеют более сложное строение мембранного комплекса.
В общих чертах картину участия ацетилхолина в осуществлении передачи нервного взбуждения можно представить следующим образом. В синаптических нервных окончаниях имеются пузырьки (везикулы) диаметром 30-80 нм, которые содержат нейромедиаторы. Эти пузырьки покрыты оболочкой, которая образована белком клатрином (отн. мол. м. 180000). В холинергических синапсах каждый пузырек диаметром 80 нм содержит ~40 000 молекул ацетилхолина. При возбуждении высвобождение медиатора идет "квантами", т. е. путем полного опорожнения каждого отдельного пузырька. В нормальных условиях под влиянием сильного импульса выделяется примерно 100-200 квантов медиатора - количество, достаточное для инициирования потенциала действия в постсинаптическом нейроне. Происходит это, по-видимому, так: деполяризация мембраны синаптических окончаний вызывает быстрый ток ионов кальция в клетку. Временное увеличение внутриклеточной концентрации ионов кальция стимулирует слияние мембраны синаптических пузырьков с плазматической мембраной и таким образом запускает процесс высвобождения их содержимого. Для выброса содержимого одного пузырька требуется примерно четыре иона кальция. Выделенный в синаптическую щель ацетилхолин вступает во взаимодействие с белком-хеморецептором, входящим в состав постсинаптической мембраны. В результате изменяется проницаемость мембраны - резко увеличивается ее пропускная способность для ионов натрия. Взаимодействие между рецептором и медиатором запускает ряд реакций, заставляющих постсинаптическую нервную клетку или эффекторную клетку выполнять свою специфическую функцию. После выделения медиатора должна наступить фаза его быстрой инактивации или удаления, чтобы подготовить синапс к восприятию нового импульса. В холинергических синапсах это происходит двумя путями. Прежде всего ацетилхолин подвергается ферментативному гидролизу. Второй путь - это энергозависимый активный транспорт ацетилхолина в нейрон, где он накапливается для последующего повторного использования.
Гидролитический распад ацетилхолина на уксусную кислоту и холин катализируется ферментом, который получил название ацетилхолинэстеразы:

В большинстве отделов головного мозга гидролиз ацетилхолина осуществляется ацетилхолинэстеразой (истинной холинэстеразой, которая гидролизует ацетилхолин быстрее, чем иные эфиры холина). В нервной ткани существуют и другие эстеразы, которые способны гидролизовать ацетилхолин, но значительно медленнее, чем, например, бутирилхолин. Эти эстеразы называются холинэстеразой (или псевдохолинэстеразой). К числу холинергических систем относятся моторные нейроны, образующие нервно-мышечные соединения, все преганглионарные нейроны автономной нервной системы и постганглионарные нейроны парасимпатической нервной системы. Большое количество холинергических симпатических областей обнаружено также в головном мозге. В зависимости от чувствительности к той или иной группе химических соединений холинергические нейроны делятся ка "мускариновые" (активируемые мускарином) и "никотиновые" (активируемые никотином). Мускариновые рецепторы ацетилхолина, имеющиеся во многих нейронах автономной системы, специфически блокируются атропином. Никотиновые синапсы присутствуют в ганглиях и скелетных мышцах. Их ингибиторами являются кураре и активный компонент этого яда - D-тубокурарин.
Необходимо подчеркнуть, что в адренореактивной системе существует два вида рецепторов для норадреналина: α- и β-адренергические рецепторы. Эти рецепторы можно отличить друг от друга по специфическим реакциям, которые они вызывают, а также по тем специфическим агентам, которые способны блокировать данные реакции.
β-Адренергические рецепторы включают эфферентную клетку с помощью аденозин-3',5'-монофосфата, или цАМФ - универсального "второго посредника" между гормонами и различными функциями клеток, на которые воздействуют гормоны.
Установлено, что как только β-адренергический рецептор (расположенный на наружной поверхности мембраны эффекторной клетки) начнет взаимодействовать с норадреналином, на внутренней поверхности клеточной мембраны активируется фермент аденилатциклаза. Затем в клетке аденилатциклаза превращает АТФ в цАМФ; последний в свою очередь способен оказывать влияние на метаболизм клетки. Этот сложный ряд последовательных реакций может быть заблокирован пропранололом - веществом, препятствующим связыванию норадреналина с β-адренергическим рецептором. Существует мнение, что β-адренергический рецептор есть не что иное, как рецепторная субъединица аденилатциклазы.
Известно, чтo в метаболизме катехоламиновых медиаторов особая роль принадлежит ферменту моноаминоксидазе (МАО). Этот фермент удаляет аминогруппу (-NH2) у норадреналина, серотонина, дофамина и адреналина, тем самым инактивируя упомянутые медиаторы. Однако в последние годы было показано, что, помимо ферментативного превращения, существует и другой механизм быстрой инактивации, точнее удаления, медиаторов. Оказалось, что норадреналин быстро исчезает из синаптической щели в результате вторичного поглощения симпатическими нервами; вновь оказавшись в нервном волокне, медиатор, естестаенно, не может воздействовать на постсинаптические клетки. Конкретный механизм этого явления пока не вполне ясен.
Адренергическая и холинергическая системы головного мозга тесно взаимодействуют с другими системами мозга, в частности с серотонической системой. В основном серотонинсодержащие нейроны сосредоточены в ядрах мозгового ствола. Нейромедиаторная роль серотонина осуществляется в результате взаимодействия серотонина со специфическими серотонинергическими рецепторами.
Исследования, проведенные с ингибитором синтеза серотонина - п-хлорфенилаланином, а также с другими ингибиторами, дают основания считать, что серотонин влияет на процессы сна. Выявлено также, что торможение кортикостероидами секреторной активности гипофиза оказывается менее эффективным у тех животных, мозг которых беднее серотонином.
Важным нейромедиатором, выполняющим тормозные функции, является γ-аминомасляная кислота (ГАМК). Количество ГАМК в головном мозге во много раз выше, чем других нейромедиаторов. Так, в гипоталамусе суммарное содержание ацетилхолина, норадреналина, дофамина и серотонина не превышает 10 мкг/г, в то время как ГАМК в этом отделе головного мозга более 600 мкг/г.
В настоящее время в терапевтической практике применяется большое количество лекарственных средств, которые действуют через систему медиаторов. Многие лекарственные вещества, успешно применяемые при лечении гипертонии, влияют на накопление и выделение адренергических медиаторов. Например, резерпин - понижающее артериальное давление средство, специфически тормозит процесс переноса катехоламинов в специальные гранулы нейронов и тем самым делает эти амины доступными действию эндогенной МАО.
Гипотензивные лекарственные препараты, такие, как α-метилдофа, под действием содержащихся в нервной клетке (аксоне) ферментов превращаются в вещества, напоминающие по своему строению норадреналин. Это "ложные" медиаторы накапливаются и выделяются вместе с естественными медиаторами, разбавляя их и тем самым снижая их эффект.
Многие антидепрессанты (вещества, снимающие депрессию) увеличивают содержание катехоламинов в синаптической щели, т. е. количество медиатора для стимулирования рецептора возрастает. К таким веществам, в частности, относятся имипрамин (блокирует поглощение норадреналина нервными волокнами), амфетамин (одновременно способствует выделению норадреналина и блокирует его поглощение), ингибиторы МАО (подавляют метаболизм катехоламинов) и др. В связи с этим возникла катехоламиновая гипотеза депрессивных состояний, согласно которой психическая депрессия связана с недостатком катехоламинов в мозге.
В начале 50-х годов фармакологи выяснили, что известный галлюциноген - диэтиламин лизергиновой кислоты (ЛСД) не только сходен по химическому строению с серотонином, но и нейтрализует некоторые его фармакологические эффекты (блокируя рецепторы серотонина). Поэтому было высказано предположение, что нарушение обмена серотонина может быть причиной возникновения особых психических заболеваний.
Считают, что такие антипсихотические средства, как аминазин (хлорпромазин) и галоперидол, усиливая синтез катехоламинов, способны блокировать дофаминовые рецепторы в мозге.
МЕХАНИЗМЫ ПАМЯТИ
ПЕПТИДЫ И БОЛЕВЫЕ РЕАКЦИИ
В 70-х годах в головном мозге различных позвоночных животных были обнаружены специфические рецепторы морфина. Эти рецепторы сосредоточены на синаптических мембранах, наиболее богата ими лимбическая система, от которой зависит эмоциональный ответ. В дальнейшем из мозговой ткани выделили эндогенные пептиды, имитирующие при инъекциях различные эффекты морфина. Эти пептиды, обладающие способностью специфически связываться с опиатными рецепторами, получили название эндорфинов и энкефалинов.
Оказалось, что пептиды с морфиноподобной активностью являются производными β-липотропного гормона гипофиза.Установлено, что β-эндорфин представляет собой фрагмент β-липотропина с 61-го по 91-й, γ-эндорфин- с 61-го по 77-й и α-эндорфин - с 61-го по 76-й аминокислотный остаток.
Энкефалины - также фрагменты β-липотропина, но они значительно меньше, чем эндорфины. Энкефалины являются пентапептидами. Наиболее изучены два пентапептида: метионинэнкефалин (Тир-Гли-Гли-Фен-Мет) и лейцинэнкефалин (Тир-Гли-Гли-Фен-Лей). Содержание метионинэнкефалинов в головном мозге в 4 раза превышает содержание лейцинэнкефалинов.
СПИННОМОЗГОВАЯ ЖИДКОСТЬ
Обший объем спинномозговой жидкости (ликвора) в норме у взрослого человека составляет около 125 мл, который каждые 3-4 ч обновляется. Ликвор рассматривают иногда как первичный транссудат или ультрафильтрат плазмы. Состав спинномозговой жидкости существенно отличается от состава плазмы крови, что и позволяет приписывать сосудистому эндотелию в нервной системе главную роль в осуществлении барьерной функции. Вода в ликворе составляет 99%, на долю плотного остатка приходится около 1%. Химический состав ликвора представлен в табл. 58.
Таблица 58. Химический состав спинномозговой жидкости
Компоненты
Содержание
Белки
0,15-0,40 г/л
Альбумины/глобулины
4:1
Остаточный азот:
8,57-14,28 ммоль/л
аминокислот
1,14-1,93 ммоль/л
мочевины
2,86-7,14 ммоль/л
Глюкоза
2,50-4,16 ммоль/л
Молочная кислота
1,67 ммоль/л
Холестерин
2,62-5,20 ммоль/л
Нейтральные жиры
Следы
Лецитин
Следы
Na+
146 ммоль/л
К+
3,5-4,0 ммоль/л
Са2+
1,5 ммоль/л
Сl-
125 ммоль/л
НСО-3
25 ммоль/л
Содержание белка в ликворе незначительно (0,15-0,40 г/л), причем отношение альбумины/глобулины равно 4; липидов в сотни раз меньше, чем в плазме крови. Возможно, что липидов плазмы крови в ликворе вообще нет. Общее содержание низкомолекулярных азотсодержащих веществ, особенно аминокислот, в 2-2,5 раза ниже, чем в крови. В ткани мозга, как уже отмечалось, количество свободных аминокислот высоко и превышает во много раз концентрацию их в крови и тем более в ликворе. Установлено, что некоторые аминокислоты (например, глутаминовая кислота) почти не проникают через гематоэнцефалический барьер. В то же время амиды аминокислот (в частности, глутамин) легко преодолевают этот барьер. Содержание глюкозы в спинномозговой жидкости относительно велико (2,50-4,16 ммоль/л), но несколько меньше, чем в крови, причем концентрация глюкозы в ликворе может повышаться или снижаться в зависимости от изменений содержания глюкозы в крови.
По содержанию натрия и калия ликвор практически не отличается от плазмы крови. Кальция в спинномозговой жидкости почти в 2 раза меньше, чем в плазме крови. Содержание хлора заметно выше, а концентрация ионов бикарбоната несколько ниже в ликворе, чем в плазме. Таким образом, минеральный состав спинномозговой жидкости также имеет характерные особенности по сравнению с таковым в плазме крови. Все это дает основание считать, что проникновение веществ через мембрану сосудистого эндотелия нервной системы - активный биохимический процесс. Источниками энергии для активного транспорта служат процесс аэробного окисления глюкозы и лишь в незначительной степени гликолиз.
Исследование спинномозговой жидкости при патологических состояниях имеет важное клиническое значение. Установлено, что при остром гнойном менингите содержание белка в ликворе может резко повышаться (5-20 г/л) по сравнению с нормой (0,15-0,40 г/л). Концентрация глюкозы в ликворе также существенно изменяется. Гипогликорахия (снижение содержания сахара в ликворе) характерна для менингита, тогда как гипергликорахия- (повышение содержания сахара в спинномозговой жидкости) наблюдается при энцефалитах, диабете и т. д. Характерно снижение концентрации хлора в ликворе при менингитах и повышение содержания его при энцефалитах. Показано также, что при менингитах, инсультах, опухолях мозга, травмах в спинномозговой жидкости повышается активность аспартатаминотрансферазы, лактатдегидрогеназы и ряда других ферментов.