- Преподавателю
- Биология
- Статья по орнитологии на тему: Температура инкубации и влажность в гнездах птиц
Статья по орнитологии на тему: Температура инкубации и влажность в гнездах птиц
| Раздел | Биология |
| Класс | - |
| Тип | Другие методич. материалы |
| Автор | Фукалов К.А. |
| Дата | 28.08.2015 |
| Формат | doc |
| Изображения | Есть |

УДК:
Фукалов Константин Александрович
Учитель биологии и химии
МАОУ "СОШ №52"
Муниципальное автономное общеобразовательное учреждение
"Средняя общеобразовательная школа №52"
614018 Россия, Пермский край, г. Пермь ул. Днепровская, 32,
e-mail: [email protected]
Температура инкубации и влажность в гнездах птиц.
Konstantin A. Fukalov
The teacher of biology and chemistry
MAEI " Secondary school №52"
Municipal autonomous educational institution
" Secondary school №52"
614018 Russia, Perm Region , Perm, Russia ul. Dnieper , 32 ,
e-mail: [email protected]
Incubation temperature and humidity of bird nests.
Аннотация. Исследованиям насиживания и инкубации у дроздов в Кишерском района, Пермского края. Были исследованы кладки дрозда рябинника и дрозда белобровика. Приводится данные исследования насиживания и инкубации у дроздов в (май - июнь) 2014-2015 г.г.. Температура инкубации, как в зоне контакта кладки с телом насиживающей птицы, так и на дне лотка гнезда очень изменчива. Птица оказывает влияние на влажность в гнезде, так как кожа насиживающей птицы обычно более влажная, чем окружающий воздух. Ключевые слова: инкубация, влажности, Пермский край, Кишерский район.
Annotation. Abstract. The subject of this research is brooding and incubation of thrushes in Kishert district, Perm region. We studied the nests of the snowbird and the redwing and here you can find the data of the researches which were undertaken in May and June, 2015. The temperature of incubation in the place of a contact between eggs and the body of a nesting bird as well as at the bottom of a nest is very changeable. The bird influences the humidity in the nest because a skin of a nesting bird is usually more humid than the air around. In Kishert district the snowbird predominates over the redwing in number.
Развитие эмбриона нельзя рассматривать вне зависимости от внешних условий. И в яйцеводе птицы или под наседкой яйцо неразрывно связанно с внешней средой, и только при определенном её состоянии возможно развитие зародыша. [2]
Говоря о среде эмбрионального развития, необходимо различать условия, непосредственно окружающие зародыш, создающиеся внутри яйца, и условия внешние по отношению к яйцу. Первые определяются наследственностью птицы, строением и составом яйца, то есть всем тем, от чего зависят его инкубационные качества. Вторые, если не считать проходящих в яйцеводе самых ранних стадиях развития, создаются в процессе инкубации.
Между внешней и внутренней средой есть определённая взаимосвязь. Изменения в яйце во время развития зародыша строго последовательны и закономерны, но смогут проявиться лишь при необходимом обогреве яйца, при определённом составе и влажности воздуха, наконец, при соответствующем положении яйца. В свою очередь, само яйцо изменяет внешнюю воздушную среду, выделяя в неё тепловую энергию, водяные пары и газы.
Сложная цепь разнообразных поведенческих реакций, закономерно сменяющих и обусловливающих друг друга на протяжении периода гнездования, составляет одну из наиболее ярких сторон видового стереотипа поведения птиц. Общий характер стереотипа поведения каждого вида в этот период эволюционно складывался под влиянием многих экологических факторов, среди которых необходимость создание и поддержания определенного температурного режима в гнезде на фоне свойственных местам его обитания климатических условий занимает одно из ведущих мест.
Устойчивый видовой стереотип гнездования и вместе с тем большая лабильность таких адаптивных форм поведения птиц, как выбор места для гнезда, подбор гнездового материала, постройка гнезда определенной формы и структуры, поведение насиживающих птиц, несомненно, играют большую роль в общем комплексе приспособлений птиц к обитанию в разных климатических условиях. Эти особенности приспособительного поведения приводят к тому, что средний температурный режим в гнездах птиц оказывался довольно сходным у видов, обитающих в различных климатических и экологических условиях; отличия средней температуры насиживаемых яиц у разных птиц относительно не велики. [11, 12]
Естественная инкубация яиц у всех без исключения птиц начиналась с откладывания первого яйца и протекала при нестабильной температуре. В большинстве случаев ее уровень в зоне контакта составлял 37-38°С при колебаниях в диапазоне 30-42°С. Колебания температуры обусловлены слетами птицы с гнезда, возникают они и при переворачивании и ориентировании яиц.
Зависимость температуры ЗК гнезда от температуры внешнего воздуха в период яйцекладки значительная и положительная, а в период собственно насиживания яиц или слабая положительная, или отрицательная.
Насиживание в период вылупления птенцов характеризовалась снижением температуры на 2-3°С, большим его колебанием и усилением тактильного фактора. Все это, по-видимому, стимулировало процесс асинхронного вылупления птенцов. [14, 13]
Относительная влажность воздуха в Пермской области во время размножения дроздов характеризуется большими колебаниями, даже в течение одних суток. Особенно резкие спады ее происходят в сухие, жаркие дни. Так, в конце апреля 1972 года минимальная влажность воздуха опускалась до 23, в мае - до 18%, т. е. она была ниже норм, рекомендуемых при искусственной инкубации. Из практики искусственной инкубации яиц известно, что как низкая (40%), так и высокая (70%) влажность воздуха вызывает изменение темпов роста зародышей и увеличивает их смертность.
Ученые считают, что на относительную влажность в гнезде не влияют ни испаряющаяся из яиц влага, ни насиживающая птица. По ее мнению, в гнезде не может поддерживаться влажность, отличная от влажности окружающего воздуха, поскольку гнездо не является хоть сколько-нибудь замкнутой системой. А.М. Болотников и С. С. Калинин (1974) нашли, что такие различия у чаек существуют. В.Д. Ефремов и В.А. Паевский (1973), Т.С. Пономарева (1974) предполагают, что определенное влияние на влажность в гнезде оказывает наседное пятно птицы на третьей стадии его развития.
Таким образом, за период яйцекладки и период собственно насиживания гнезда рябинников теряют около 50% веса за счет испарения свободной воды. Это позволяет предполагать, что строительный материал может оказывать влияние на микроклимат гнезда, стабилизируя режим влажности, по крайней мере, в период яйцекладки и первую половину собственно насиживания. Такое приспособление дает возможность виду успешно размножаться в условиях колеблющейся влажности воздуха и расширяет во времени сроки гнездования. [7]
В мае был составлен и проложен маршрут с целью учёта количества гнёзд дрозда-белобровика и дрозда-рябинника. Маршрут проходил в Кишертском районе вдоль берега реки Лёк, между деревнями Киселёво - Медведево, и по протяжённости составил порядка трёх километров.
Помимо учёта количества гнёзд измерялись и метрические показатели гнёзд обоих видов: высота расположения гнезда, высота самого гнезда, ширина гнезда, его глубина. Высота расположения гнезда измерялась непосредственно от земли метром с точностью до 20см. Высота гнезда, ширина гнезда и его глубина измерялись линейкой с точностью до 5мм. Вдобавок нами было зафиксировано, на каком дереве и в какой его части располагается гнездо.
Также нами проводились измерения метрических показателей яиц дроздов обоих видов: длинна и ширина яиц. Измерения проводились штангенциркулем с точностью до 0,1мм.
Измерялись все гнезда, попавшие нам на протяжении маршрута, кроме тех, до которых было невозможно добраться технически. Всего было измерено 17 гнёзд дрозда рябинника и 6 гнёзд дрозда белобровика. Из-за сроков проведения измерений в некоторых гнездах были обнаружены птенцы, в таких гнёздах измерялось только само гнездо, птенцы измерению не подвергались.
По завершению исследования полученные данные статистически обрабатывались, результаты которых описаны ниже.
В течение полевых сезонов (май - июнь) 2014 - 2015г.г. изучена температура и влажность в гнёздах дрозда белобровика и рябинника. Применялся регистрирующий комплекс iBDL-HS (рис. 1). Комплекс состоит из логгеров - таблеток и считывающего устройства для размещения информации в компьютере. Время работы логгера зависит от частоты внесения информации в его энергонезависимую память. В каждое гнездо помещали два логгера, один из которых в зону контакта яиц с наседным пятном птицы (в дальнейшем - ЗК), второй на дно лотка под кладку. Период внесения информации в память - 25 сек. Третий логгер обеспечивал регистрацию внешней температуры непосредственно у гнезда.

(Рис.1. Регистрирующий комплекс iBDL-HS.)
За температуру инкубации мы принимаем температуру ЗК, то есть практически температуру наседного пятна насиживающей птицы. Регистрация температуры дна лотка гнезда позволяет выяснить возникающий термоградиент в гнезде, силу влияния, как температуры зоны контакта, так и внешней температуры на развивающийся эмбрион и оценить силу их связи. [9]
Математическую обработку полученного материала проводили по общепринятым методикам. За показатель корреляции мы брали не коэффициент, так как он используется тогда, когда связь между признаками линейна, а в нашем случае она оказалась не линейна. Поэтому мы воспользовались корреляционным отношением - удобным показателем силы связи признаков. Для вычисления силы влияния одного параметра на другой, например одной температуры на другую, мы применяли однофакторный дисперсионный анализ. [8]
На протяжении маршрута в 3 км нами было измерено 17 гнёзд дрозда рябинника и 6 гнёзд дрозда белобровика. Таким образом, соотношение оказалось равным приблизительно 3 : 1.
Оба вида в основном предпочитали, для строительства гнёзд, либо ольху, либо сухой вяз мелколистный. В большинстве случаев гнездо располагалось в развилке стволов, либо в развилке веток, иногда во впадине ствола сухих сломанных деревьев. Различающихся предпочтений в выборе места строительства гнёзд у разных видов не наблюдалось. При сходных условиях существования, одинаковой кормовой базе рябинник более доминантен, чем белобровик. Исходя из этого, заметно, что на территории Кишертского района, вдоль берега реки Лёк, между деревнями Киселёво и Медведево численно преобладает дрозд-рябинник по сравнению с белобровиком.
Измерялись и метрические показатели гнёзд обоих видов: высота расположения гнезда на дереве, высота самого гнезда, ширина гнезда, его глубина. Первый показатель был измерен с точностью до 20 см, последующие с точностью до 5 мм.
Средняя высота расположения гнезда рябинника составила 3,5±1,24 м над землёй, а белобровика 5,17 ± 0,41, то есть белобровик предпочитает строить гнезда выше, чем рябинник примерно на 1,6 м. Возможно, это связано с меньшей численности белобровика в биотопе.
Средние размеры. У дрозда-рябинника высота гнезда составила: 11,4±0.65 см, ширина гнезда - 14,8±0.98 см и глубина гнезда - 5,4±0.28 см. У белобровика высота гнезда составила: 11,3±0.61 см, ширина гнезда - 15±0.55 см и глубина гнезда - 5,47±0.29 см. Разница в размерах гнёзд между видами - незначительна: 0,07, 0,2 и 0,07см, соответственно. Сходство в размерах, форме и месте расположения гнезд вероятнее всего можно объяснить близким систематическим родством (принадлежностью к одному роду Turdus).
Так же можно отметить взаимосвязь высоты и ширины гнезда (рис.2)

(Рис.2. Взаимосвязь высоты гнезда и его ширины у дроздов белобровика и рябинника.)
На рис. 2 наглядно видно, что высота гнезда обратно пропорциональна ширине гнезда, т.е. при отклонении от средней высоты, так же отклоняется ширина. Особенно хорошо это прослеживается на гнездах №№: 2, 5, 7, 8, 16, 19, 22, 24.
На протяжении нашего маршрута нами было измерено 93 яйца дрозда рябинника и 36 яиц дрозда белобровика. Количество яиц в гнезде варьировало от 4 до 8 у рябинника, и от 4 до 5 у белобровика. Среднее количество яиц гнезде у рябинника составило 5,5, а у белобровика 6,0. Мы измеряли метрические показатели яиц дроздов обоих видов: длину и ширину яиц. Измерения проводились штангенциркулем с точностью до 0,1мм. Средняя длина яиц рябинника составила 26,18±0,14мм, а средняя длина яиц белобровика - 25,61±0,1 мм. Средняя ширина яиц рябинника составила 19,12±0,10мм, а средняя ширина яиц белобровика - 18,85±0,10мм. Разница в длине яиц составила 0.57мм, а разница в ширине составила 0,55мм.
Исходя из этих данных, можно сказать, что яйца дрозда-рябинника крупнее яиц белобровика на 0.57мм в длину, и а 0.55мм в ширину. Но такая разница составляет порядка 2%. Исходя из этого, можно сказать, что яйца дрозда-рябинника и яйца дрозда белобровика имеют схожие размеры. Сходство в размерах яиц вероятнее всего можно объяснить так же близким систематическим родством (принадлежностью к одному роду Turdus).
Температура в зоне контакта исследована у дроздов рябинника и белобровика. Температура ЗК - самая высокая в гнезде, так как по сути, это температура наседного пятна птицы. Главная особенность этой температуры - её нестабильность. Малейшее шевеление наседки, не говоря уже о сходах её с гнезда, отражаются на уровне этой температуры (рис.3).
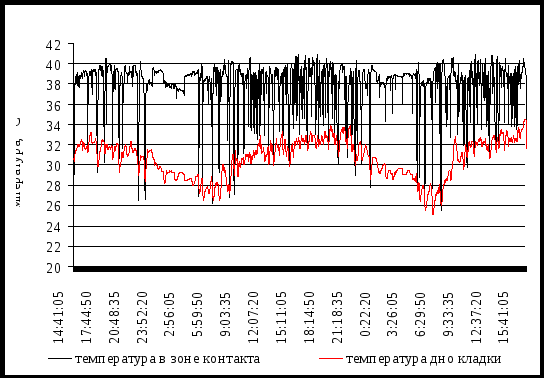
(Рис. 3. Температура в гнезде дрозда-белобровика на восьмые и девятые сутки собственно насиживания.)
Множество направленных вниз пиков на температурной кривой информирует о постоянных движениях насиживающей птицы, а так же многократных сходах с гнезда. Таких уходов за двое суток инкубации изображённых на рис.4 мы насчитали 39 в дневное время. Во время сходов птицы с кладки температура начинает быстро снижаться. Эти понижения могут достигать 25-26˚С по сравнению с общим уровнем температуры ЗК в 38-39˚С, то есть примерно на 13˚C. Общий размах колебаний температуры в ЗК от минимального до максимального значения составил 26,9˚C у рябинника и 20,5˚C у белобровика. Температура на дне лотка колеблется ещё больше: у рябинника в пределах 28,7˚C, у белобровика - 24,3˚C.
Согласно визуальному наблюдению из укрытия и с помощью беспроводной камеры слежения установленной в 1метре от гнезда самка проводит в гнезде подавляющее время - примерно 90% дневного, а ночью всегда. Дневная активность, в виде утреннего ухода с гнезда начиналась в течение восьми регистрируемых суток в 4:08-4:45 и заканчивалась в 22:42-23:57, причём в пасмурные дни «ночь» начиналась несколько раньше, примерно на 30-40 мин. а «день» - позже. Таким образом, «день» для рябинника и белобровика длится почти 20 часов, тогда как «ночь» - всего 4.
У дроздов рябинника и белобровика практически одинаковое среднеарифметическое значение температуры инкубации (ЗК) (табл.3). Она менее стабильна, чем температура на дне лотка гнезда, о чём говорит большее значение среднего квадратического отклонения у неё. Сила связи этих температур очень высока и близка к функциональной. То же самое можно сказать и о силе влияния температуры ЗК на температуру дна.
Температура в гнёздах мало зависима от внешней температуры. Эта зависимость несколько больше у температуры дна и совсем мала у температуры ЗК. Казалось должно быть наоборот - уходы птицы ведут к мгновенному снижению температуры в ЗК, а температура дна лотка более инертна. Тем не менее, при экстремальных значениях внешней температуры в дневную жару или в ночной холод наседка старается не сходить с гнезда и остаётся здесь на большее время. Таким образом, относительная независимость температурного режима гнезда обеспечивается поведением наседки (плотностью насиживания).
Поскольку температуры на верхнем уровне отложенных яиц и на дне лотка отличаются друг от друга, то в гнезде возникает вертикальный термоградиент обусловленный, как правило, размерами яиц (в основном их малым диаметром). Согласно полученным данным, он несколько больше у рябинника и меньше у белобровика.
-
Болотников А.М. Соколова Т.И. Чащин С.П. Циклы яйцекладки плодовитость и выживаемость птенцов грача в Пермской области /Сб. статей: Ученные записки / Перм. гос. пед. ин-т. Пермь. 1973. Т. 113. С. 39 - 41.
-
Болотников А.М., Шураков А.И. Каменский Ю.Н., Добринский экологическое равновесие онтогенеза птиц. - Сверловск: УНЦ АН СССР 1969 - 228 с
-
Болотников, А.М. Экология раннего онтогенеза птиц // А.М. Болотников. - Свердловск: УНЦ АН СССР, 1985
-
Гражданкин А.В. Исследования гнездовой биологии птиц и моделирование гнездовых условий // Актуальные проблемы орнитологии. « Наука». М. 1984. С. 217 - 228
-
Данилов Н.Н., Шварц С.С. Экологическая физиология птиц и некоторые перспективы ее развития. // Н.Н. Данилов, С.С. Шварц Орнитология в СССР. - Ашхабад, 1969, кн. 1, - С. 89-99.
-
Дементьев В.Г., Гладков Н.А. Птицы Советского Союза. - Москва: Изд-во «Просвещение» 1954. С. 263 - 265.
-
Евдокимов В.Д. О величине кладки и размерах яиц рябинника в разных биотопах// Гнездовая жизнь птиц. - Пермь: ПГПИ, 1980. С. 80 - 82.
-
Лакин Г.Ф. Биометрия/ Г.Ф. лакин - М.:Высш. Шк. 1980. С. 125 - 200.
-
Литвинов Н.А., Шураков С.А., Ганщук С.В., Сергеев В.В Характеристика температуры инкубации у трех видов птиц / Н.А.Литвинов, С.А. Шураков, С.В. Ганщук, В.В. Сергеев // Вестник уральской медицинской академии наук. Екатеринбург, 2009. - С. 234-235.
-
Отрыганьев Г.К. Внешняя среда эмбрионального развития// Инкубация. «Колос». М. 1984. С.88 - 101.
-
Шилов И.А. Регуляция теплообмена у птиц / И.А. Шилов - М.: Изд-во МГУ, 1968 - С. 104 - 107, 175 - 176, 252.
-
Шилов, И.А. Регуляция теплообмена у птиц // И.А. Шилов - М.: Изд-во МГУ, 1968. - С. 104-107, 175-176, 252.
-
Шкарин В.С. Размеры и форма гнёзд рябинника // Гнездовая жизнь птиц. - Пермь: ПГПИ, .С.106 - 108.
-
Шураков, А.И. Строительный материал и влажность гнезд рябинника в Пермской области // Гнездовая жизнь птиц. - Пермь: ПГПИ, 1975. С. 48-52.
9